Radiotherapy, which traditionally has been used for localized tumor treatment, is being increasingly combined with CPIs to treat metastasized tumors [49, 50]. Radiation can kill cancer cells, release tumor antigens, and subsequently activate tumor specific immune response [51]. Consequently, tremendous effort has been made to optimize radiotherapy in order to maximizing its immune-activation capacity [52, 53].
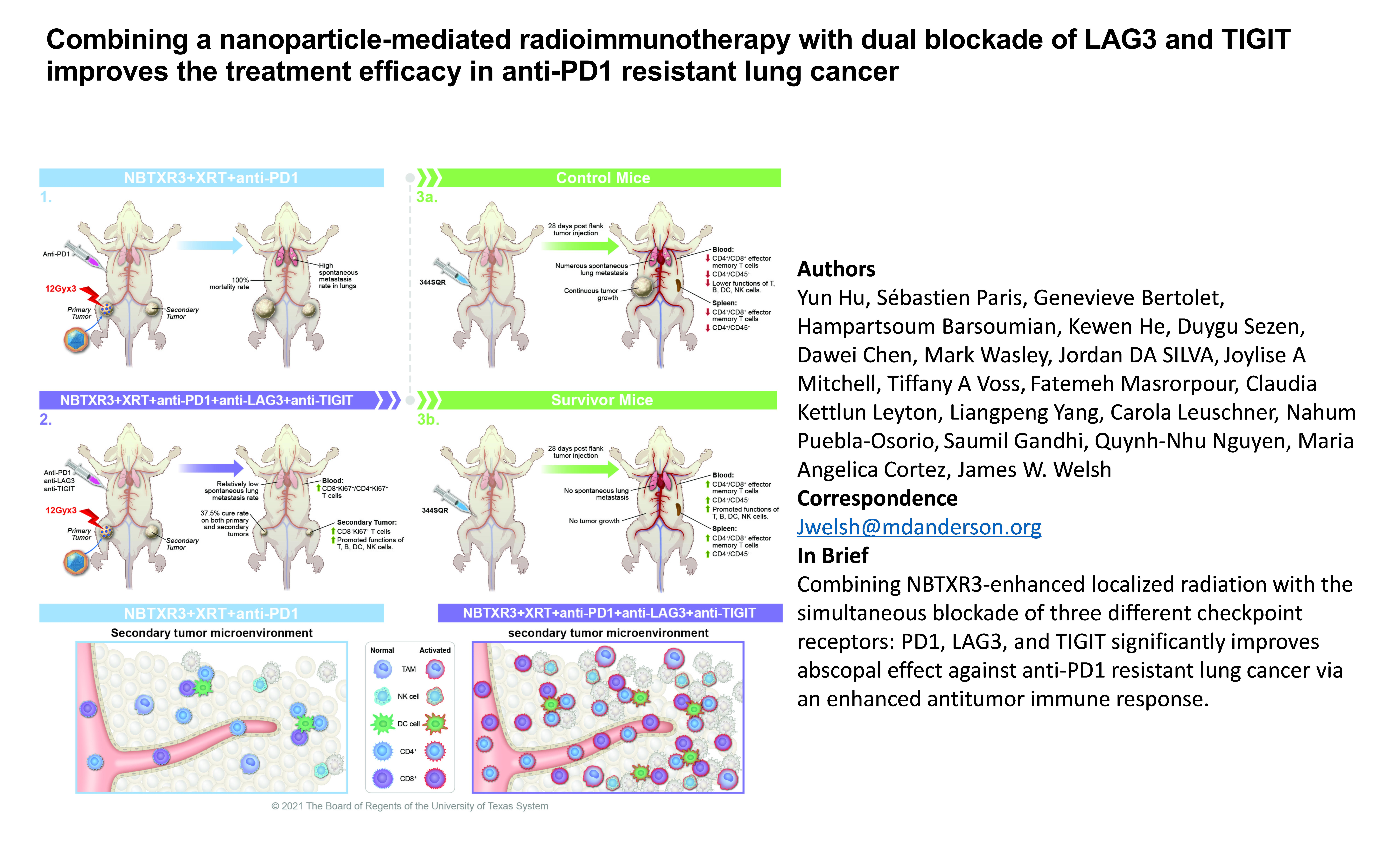
In pursuit of this end, we previously discovered that the radiation-enhancing nanoparticle NBTXR3 could significantly promote the activation of radiotherapy-mediated antitumor immunity. However, the activated effector immune cells, including CD4+ T cells, CD8+ T cells, and NK cells, are subject to exhaustion [54]. Numerous ICRs, such as PD1, LAG3, and TIGIT, have been found to participate in the exhaustion of immune cells [55]. Therefore, in this study, we explored the possibility of increasing immune activation while reducing immune cell exhaustion through combining NBTXR3-enhanced radiation with blockade of LAG3, TIGIT, and PD1.
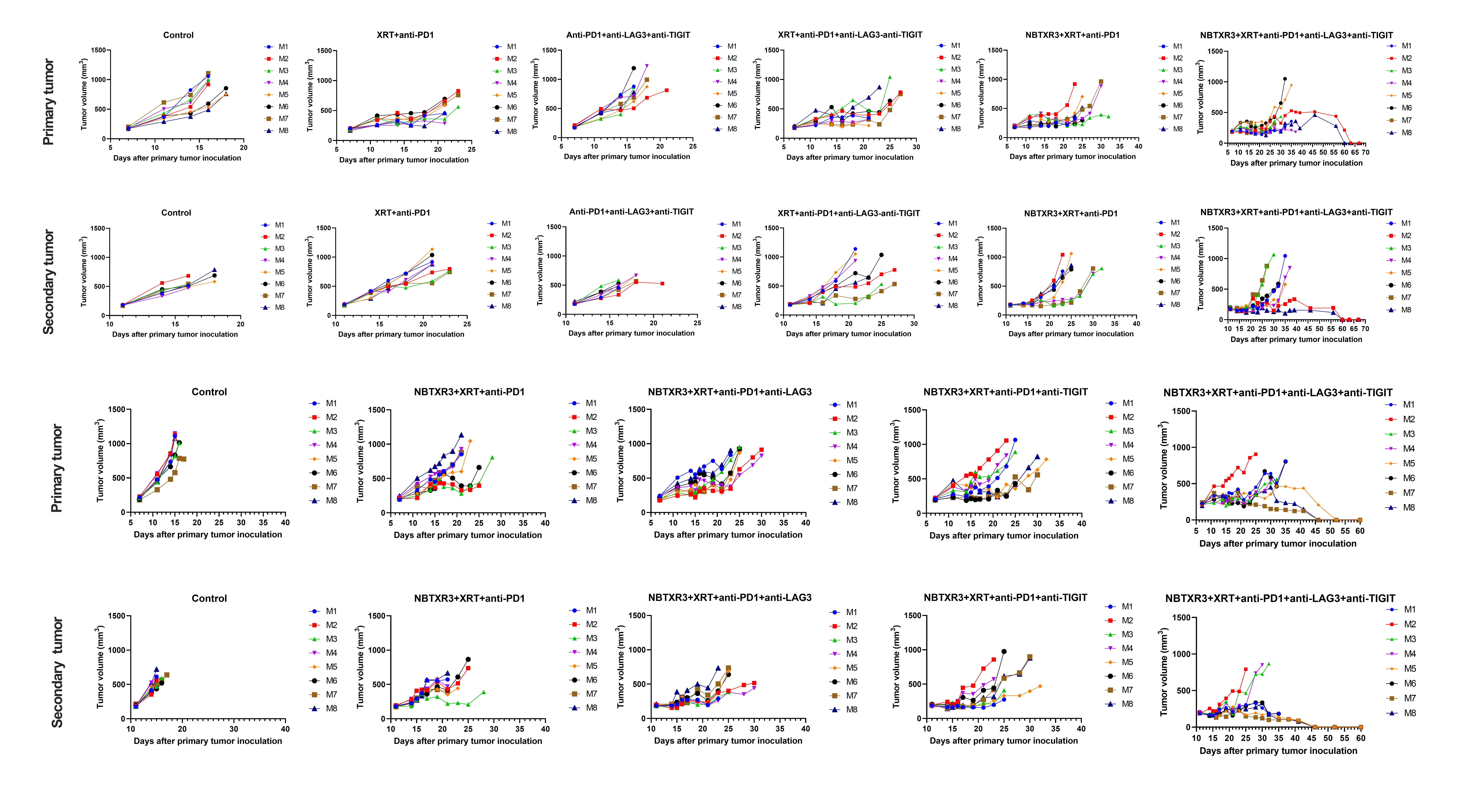
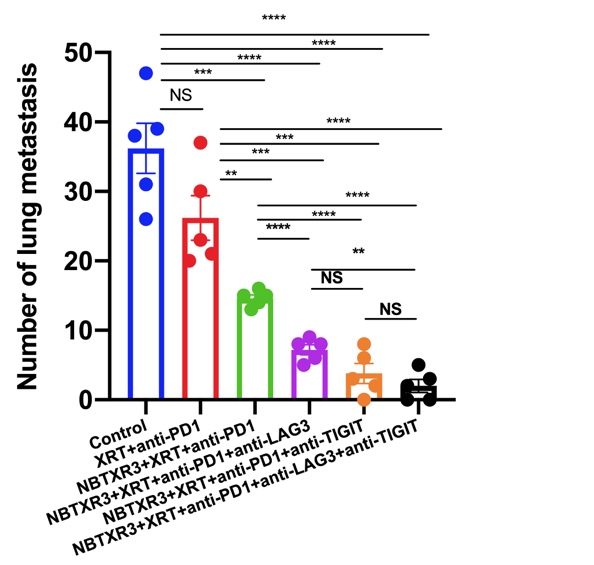
In agreement with our previous findings [20], mice treated with NBTXR3 + XRT + αPD1 displayed improved control of both primary and secondary tumors over that of XRT + αPD1. No treatment group that lacked NBTXR3 showed as robust of a therapeutic response, even with triple CPI, alone or with radiation. These results demonstrate the impressive potency of NBTXR3 in amplifying tumor clearance. We found, moreover, that adding αLAG3 or αTIGIT alone to NBTXR3 + XRT + αPD1 significantly augmented this anti-tumor effect, decreasing growth of both tumors in a manner superior to that of NBTXR3 + XRT + αPD1 without further CPI. Furthermore, inhibiting all three checkpoints simultaneously, along with NBTXR3-amplified irradiation (NBTXR3 + XRT + PLT), achieved unparalleled efficacy in tumor clearance, reduction of metastasis, induction of immunological memory, and overall survival. Indeed, the survival rate of mice treated with NBTXR3 + XRT + PLT was 37.5%, compared to 0% survival in the other groups.
There have been reports showing that the dual immunotherapies of αLAG3 + αPD1 or αTIGIT + αPD1 can effectively enhance antitumor immunity and inhibit tumor growth in other tumor models [56, 57]. However, the results of this study suggest that, in tumors already resistant to αPD1, NBTXR3-enhanced radiation is essential for this CPI-mediated immune response. We hypothesize that this is likely due to the initial radiation-induced trauma to the primary tumor causing inflammation and tumor-associated antigen exposure, effectively penetrating the immune privilege enjoyed by the tumor and kick-starting immune activation. The corollary of this hypothesis is that, just as immune activation requires NBTXR3-amplified XRT to initiate, NBTXR3-amplified XRT requires immune activation to potentiate the full therapeutic effect of the treatment. Indeed, we found that control of tumor growth and animal survival were ablated when immune cells were selectively depleted.
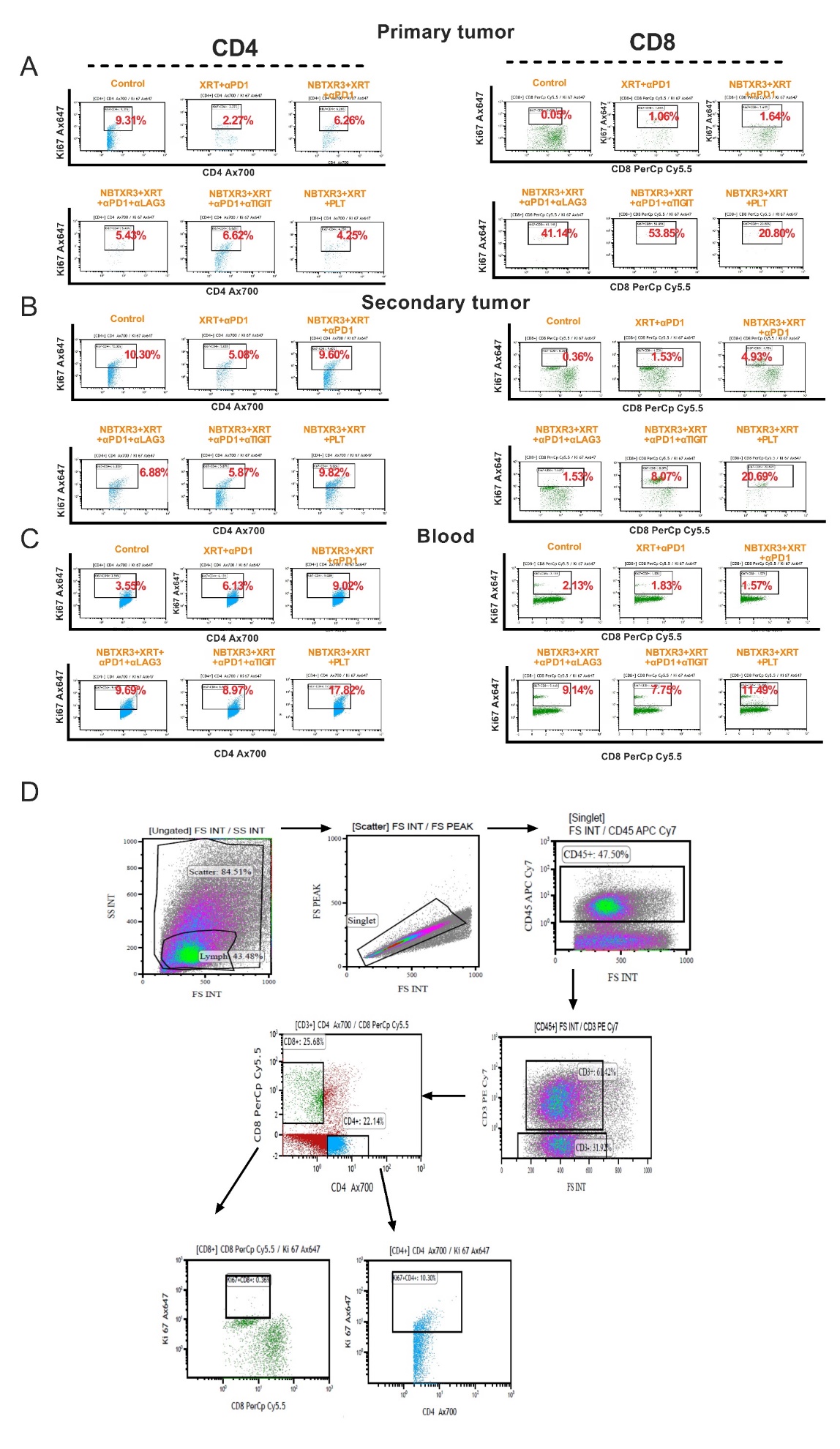
Tumor clearance is typically thought to be the domain of cytolytic immune cell populations – CD8+ T cells and natural killer (NK) cells T cells. However, the immune population whose depletion most strongly impacted the therapy’s efficacy were CD4+ T cells, the so-called helper T cells. Helper T cells serve as the generals of the immune response, directing and selectively enhancing the activities of other immune cells through the secretion of cytokines. Like generals, CD4+ T cells require information from scouts on the nature of the threat. In the immune system, this information comes to the CD4+ T cells in the form of tumor antigenic peptides presented by antigen-presenting cells (APCs), the most prominent members of which are dendritic cells (DCs) and macrophages. Macrophages are typically first on the scene of a site of inflammatory injury – such as from radiation damage to a tumor. Once there, they set to work engulfing damaged and pathogenic cells through phagocytosis, digesting the engulfed cells and presenting the digested peptides to CD4+ T cells via class II major histocompatibility complexes (MHCs). The CD4+ T cells, having thus been appraised of the threat, secrete inflammatory cytokines such as IFNγ, which stimulate the macrophages to step up their attack. They, in turn, begin secreting their own inflammatory cytokines such as IL-6 and tumor necrosis factor (TNF), producing caustic reactive oxygen species (ROSs) that seep into the tumor and induce apoptosis, and intensifying their phagocytic engulfment of diseased or damaged cells. CD4+ have been previously observed to be the primary mediators of tumor clearance in manner that is independent of CD8+ T cells but reliant on IFNγ [58], and the partnership between CD4+ T cells and macrophages has been documented as one of the mechanisms driving this effect [59].
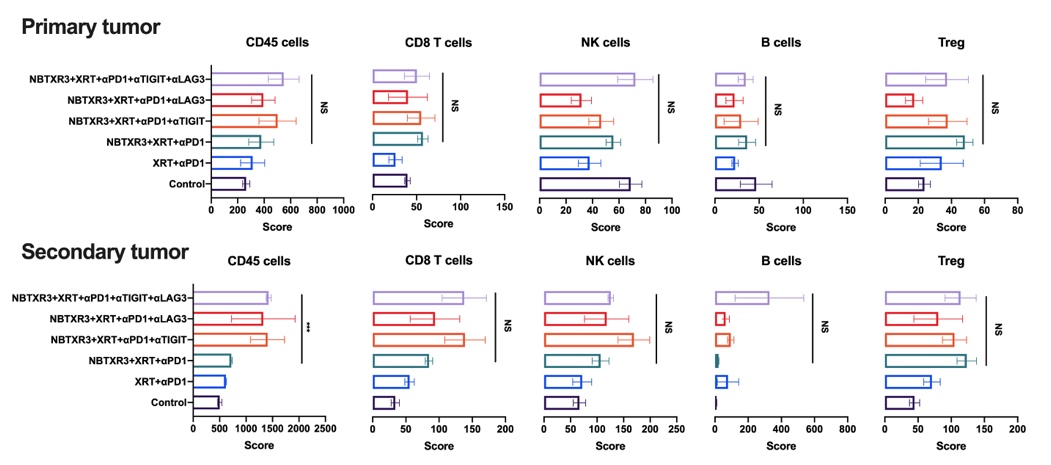
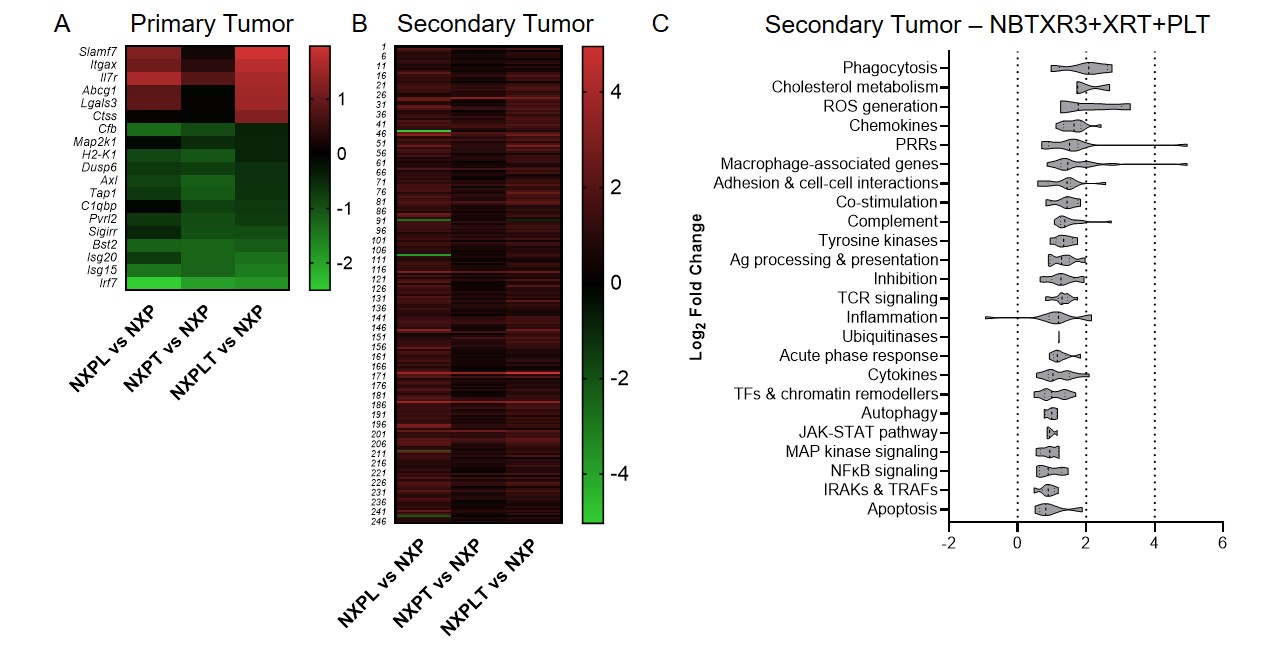
When we looked at the mRNA transcriptomic signature of primary and secondary tumors of mice treated with NBTXR3 + XRT + PLT, we observed a textbook upregulation of genes participating in every step of this process. At the primary tumor, we saw strong upregulation of genes primarily associated with macrophage activation. When we turned to the secondary tumor, we saw a full-scaled activation of the immune response. Genes associated with antigen presentation were upregulated, as were genes associated with macrophage function and identity. In accordance with this, numerous genes specifically involved in phagocytosis and ROS generation were also heightened. Furthermore, we detected the upregulation of genes associated with T cell activation – specifically, CD4+ T cell activation. Inflammatory cytokines such IFNγ and IL-6, components of their receptors, and downstream signaling intermediaries were likewise elevated. Several members of the complement system, which can be generated by both DCs and T cells, were also increased. While classically thought of as antibacterial and antiparasitic defense, the complement system can also be activated by the altered membranes of tumor cells induced to apoptotic and necrotic cells [46]. Conspicuously absent were any genes associated with CD8+ T cell-specific responses.
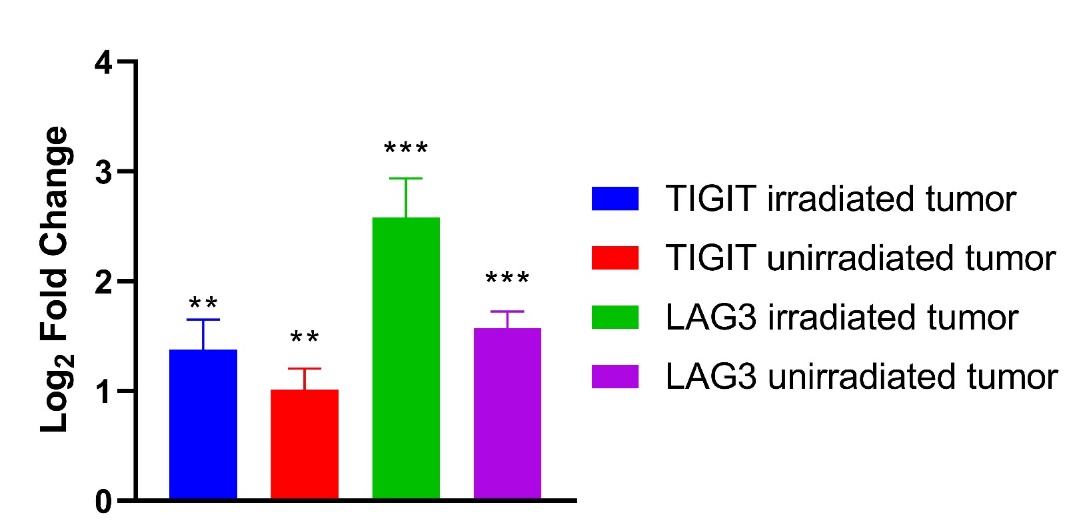
Immune-related gene upregulation in the secondary tumor was much weaker in mice treated with NBTXR3 + XRT + αPD1 and αTIGIT than in mice treated with NBTXR3 + XRT + PLT. However, it was equally robust in mice treated with NBTXR3 + XRT + αPD1 and αLAG3 as in NBTXR3 + XRT + PLT-treated mice (Fig. S5B). While control of primary tumor growth and overall survival was greater for NBTXR3 + XRT + PLT-treated mice than for mice treated with only NBTXR3 + XRT + αPD1 and αLAG3, control of the secondary tumor growth was roughly equivalent between them. These results suggest that the key difference in outcome between these two treatments stems from the control of the primary tumor. Another possibility, not exclusive to the previous, stems from the exact genes that were upregulated in the secondary tumors of mice treated with NBTXR3 + XRT + PLT versus those treated with NBTXR3 + XRT + αPD1 and αLAG3. In particular, we found that, while the gene for IFNγ was, in fact, downregulated in mice treated with NBTXR3 + XRT + αPD1 and αLAG3 (as compared to mice treated with only NBTXR3 + XRT + αPD1), it was markedly upregulated in mice treated with NBTXR3 + XRT + PLT. Also of note is that, in mice those treated with NBTXR3 + XRT + αPD1 and αLAG3, the gene for PD-L1 was expressed at a level higher than that expressed by mice treated with NBTXR3 + XRT + PLT, possibly indicating greater immune exhaustion.
Regardless of the cause, what is clear is the differential combination of αPD1, αLAG3, and αTIGIT in combination with NBTXR3-amplified irradiation achieves similar but distinct outcomes. We suspect that what might underlie the differences between these therapies is that the combination of NBTXR3-enhanced irradiation of the primary tumor, in tandem with blockade of all three ICRs, serves to push the immune response over some threshold at the primary tumor site, allowing complete or nearly complete clearance thereof. This effect is not attributable to the action of any one of these components, as only the presence of all of them together achieves this outcome. How, exactly, the elements of this combined approach harmonize in such a way as to achieve this highly favorable outcome is richly deserving of further study. What is clear, however, is that the key lies in how each lifted checkpoint influences the immune response.
The heavy dependence of the NBTXR3 + XRT + PLT therapy on the immune responses is reflected by the faster growth of both the primary and the secondary tumors after depletion of CD4+ T cells, CD8+ T cells, and NK cells. It is also noteworthy that these three types of immune populations seem to have varied influence in antitumor efficacy, in which depletion of CD4+ T cells completely abrogated the antitumor effect, while depletion of NK cells had the least impact on the antitumor activity. Other studies have also reported results that attest to the indispensability of CD4+ T cells to the efficacy of RIT [60, 61]. The critical role of CD4+ T cells may be attributed to the fact that they lie upstream of CD8+ T cell- and NK cell-mediated antitumor pathways.
The reliance of this combinatorial therapy on the immune system – and its potential for stimulating it – is perhaps no better illustrated than with the strong memory response to repeat exposure to the same tumor cells in mice that cleared the initial tumors. These mice display superior memory cell numbers and mobility to their uninoculated counterparts, effectively immunizing them against relapse, even when it is deliberately induced. This remarkable finding indicates that this line of therapy may provide patients not only with a highly favorable response, but even complete and enduring remission.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}