In this study, the molecular impacts of PRP treatment on the human endometrium in a primary culture were identified first. In the proliferative phase, the exposure of estrogen in the endometrium increases endometrial thickness and the linear growth of endometrial glands and angiogenesis, which are processes that are indispensable for pregnancy. PRP was found to strongly promote cell migration and proliferation and induce gene expressions associated with cell growth, including the proinflammatory response in undifferentiated HESCs. In another study, PRP was found to induce endometrial regeneration with cell proliferation and migration in human endometrial cell lines [27], which was in agreement with our results. PRP containing abundant growth factors, such as transforming growth factor-β, platelet-derived growth factor, insulin-like growth factor, vascular endothelial growth factor, and epidermal growth factor [28], is expected to accelerate cell growth. In addition, PRP treatment led to a considerably high expression of many histone genes (HIST1H1A, HIST1H1B, HIST1H2A, HIST1H2A1, HIST1H2AB, HIST1H2AJ, HIST1H2BM, HIST1H3B, HIST1H3F, HIST1H3G, HIST1H3J, and HIST1H4D) in undifferentiated HESCs (data not shown). An increase in acetylation levels of H2AK5, H3K9, and H4K8 is required to enable endometrial regeneration and proliferation in the early proliferative phase [29, 30]. PRP may also induce histone acetylation for cell proliferation. In fact, all previous clinical studies have shown that intrauterine PRP infusion increases endometrial thickness in infertile women with thin endometria [19–25]. According to our results, PRP has a high potential to repair impaired endometrium such as in Asherman syndrome (AS). In a murine model of AS, human PRP improved endometrial morphology and reduced fibrosis, resulting in an increased number of live births [31]. Although PRP did not considerably improve postsurgical adhesion, endometrial thickness, and the pregnancy rate after hysteroscopic adhesiolysis in some randomized clinical trials with a small sample size [32, 33], some successful pregnancy cases have been reported [34]. In our study, PRP demonstrated great potential for repairing a damaged intrauterine environment; thus, there is a need to reexamine the treatment protocol and conduct further clinical trials.
PRP also up-regulated the genes involved in the beta chain of the T-cell receptor (TRBV3-1, TRBV6-5, TRBV7-2, TRBV27, TRBC2, and TRBJ2-3), which is responsible for antigen recognition and activation of cellular immunity during bacterial infection [35]. Chronic endometritis (CE) is a representative intrauterine disorder characterized by the continuous inflammatory status of the local endometrium, which can cause implantation failure and pregnancy loss [36]. Most cases of CE are caused by intrauterine infection with a wide variety of micro-organisms; thus, the current treatment protocol for CE is broad-spectrum antibiotic therapy [37]. However, inappropriate systemic antibiotic therapy increases the risk of the emergence of antibiotic-resistant bacteria and should be avoided [38]. Intrauterine PRP infusion with antibiotic effects is expected to be used for the treatment of CE, instead of antibiotic therapy. Recently, some studies have already reported successful pregnancy outcomes after PRP infusion as a treatment for CE [39, 40]. Further clinical trials on PRP treatment for CE are warranted in the future.
PRP influenced a large number of genes in undifferentiated HESCs, whereas in decidualized HESCs, the number of the genes affected by PRP was extremely low as low as one-sixth of the undifferentiated HESCs. Cross-referencing of the array data identified only 10 genes that are regulated by PRP in undifferentiated and decidualized HESCs. Intriguingly, PRP treatment both positively and negatively affected inflammation and cell proliferation in undifferentiated and decidualized HESCs, respectively. The function and physiology of endometrial development are completely different in the secretory phase (decidual phase) than that in the proliferative phase. Decidualization starts with an acute stress response with the secretion of proinflammatory mediators [41, 42]. In the initial decidual phase, some stromal cells burdened by replication stress fail to differentiate as senescent decidual cells [41, 42]. Senescent decidual cells secrete the senescence-associated secretory phenotype, including proinflammatory cytokines and chemokines, thus inducing secondary senescence in adjacent decidual cells [42]. Uterine natural killer (uNK) cells deplete their senescent counterparts [41, 42]. After several days of inflammatory reprogramming, anti-inflammatory decidual cells emerge that are highly resistant to metabolic and oxidative stress [4, 43]. In clinical studies using ultrasonography, endometrial volume showed a constant increase as a result of an increase in estrogen level in the proliferative phase; however, the increase in endometrial thickness stopped during the secretory phase after ovulation [44]. Recent reports have shown that a decrease in endometrial thickness after ovulation (endometrial compaction) is an important finding for successful pregnancy [45, 46]. Considered together in the process of decidual change in HESCs, senescent decidual cells are eliminated and specialized anti-inflammatory decidual cells emerge with metabolic and oxidative stress resistance, leading to a slightly decreasing endometrial volume. During the secretory phase, PRP has a potential for promoting endometrial compaction with an anti-inflammatory response, which supports successful pregnancy.
Decidual transformation of the endometrium and subsequent trophoblast invasion require down-regulation of the PI3K/AKT signaling pathway [26, 47]. PI3K is composed of two subunits: the catalytic subunit, p110, referred to as PIK3C, and an adaptor/regulatory subunit, p85, also referred to as PIK3R [48]. PI3K enzymes primarily act to convert abundant cellular phosphatidylinositol-4,5-bisphosphate (PIP2) to phosphatidylinositol-3,4,5-triphosphate (PIP3), which mediates AKT phosphorylation and downstream signaling. Phosphatase and tensin homologue (PTEN) antagonizes PI3K-dependent signaling via the dephosphorylation of PIP3 to PIP2 [49]. PTEN is abundantly expressed in decidualized stromal cells compared with undifferentiated cells; therefore, the increased PTEN diminishes the effects of PI3K signaling and regulates vascular remodeling through the infiltration of uNK cells during decidualization [50]. In our results, PRP was shown to up-regulate the expression level of PTEN and decrease the expression of the main subunits of PI3K, PIK3CA (p100α), and PIK3R1 (p85α). Therefore, PRP mainly decreased the expression of PI3K by upregulating the expression of PTEN. In burn injury treatments, PRP also induced the expression of PTEN and suppressed the expression of PI3K/AKT and its related pathways, resulting in the alleviation of neuropathic pain [11]. Furthermore, some reports showed that TC2N and SERPINE2 (also known as PN1) in up-regulated genes by PRP have suppressive effects on the PI3K/AKT signaling pathway [51, 52], and RASGRP2 and SPHK1 in down-regulated genes were shown to regulate the PI3K/AKT signaling pathway [53, 54]. However, the expression of PIK3C3, also known as Vps34, among the PI3K genes increased after PRP treatment. PIK3C3 is a kinase that is indispensable for autophagy and the lysosomal degradation pathway, which regulates intracellular homeostasis and protects organisms against infection, cancer, and aging [48, 55]. Therefore, PIK3C3 does not regulate the main PI3K/AKT signaling. When PI3K signaling was inhibited by PRP in decidualized cells, AKT expression was also expected to be suppressed, but this gene was not identified in our results. In the process of decidualization, AKT phosphorylation is decreased, yet the expression of total AKT is not affected [56]; thus, AKT may not be detected in our results.
Moreover, PRP down-regulated the expression of toll-like receptor 4 (TLR4) and its downstream genes MAPK14 (also known as p38), DEP domain containing MTOR-interacting protein (DEPTOR), and Myc. TLR4 is the integral receptor in the response to protect against the non-self-antigens, including microorganism infection with inflammatory response as a Th1-dominant cell balance [57]. PI3K signaling is also regulated via TLR4 [57]. Successful pregnancy requires maternal immune tolerance to a semiallogeneic embryo with a balance of proinflammatory cytokines and antiinflammatory cytokines, secreted by Th1 and Th2 cells, respectively, in favor of a Th2 dominance [58]. RIF and recurrent pregnancy loss have been linked to an aberrantly high Th1/Th2 cell ratio [59]. An infusion of PRP may regulate high Th1 response via a decrease in the expression of TLR4 signaling and improve endometrial immune tolerance for implantation in decidualized endometrium.
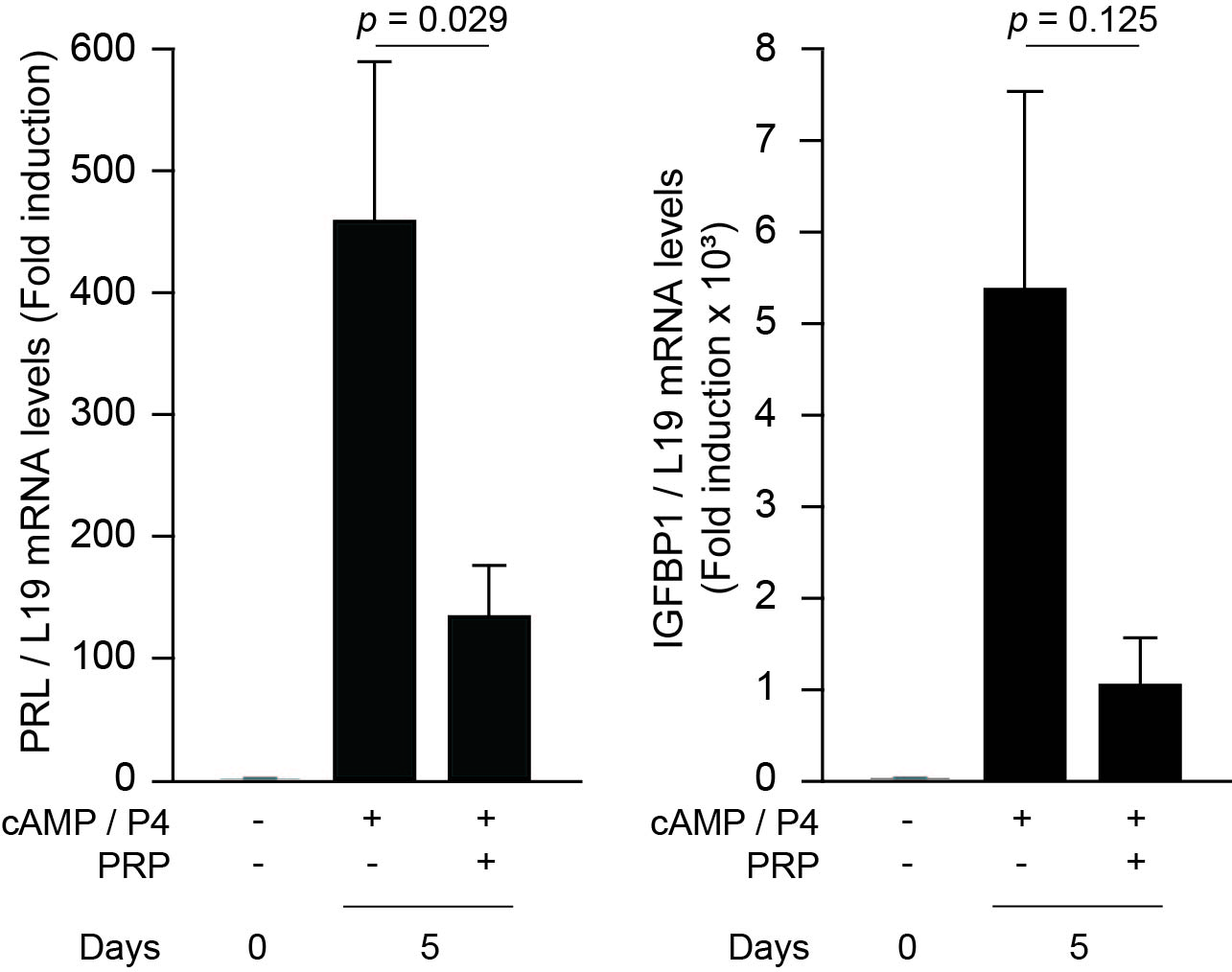
PRP treatment has many beneficial effects in terms of improving endometrial receptivity; however, PRP diminished the expression of decidual marker genes. One of the markers, IGFBP1, is predominantly produced by senescent decidual cells in primary cultures [42]. It may result from a suppressive effect of PRP on cell proliferation of senescent cells, yet PRP may inhibit the decidualization of HESCs, resulting in a detrimental effect on pregnancy outcomes in unselected women who do not have an impaired endometrial environment. In previous clinical trials that did not consider the inclusion criteria of endometrial status, the therapeutic effect of intrauterine PRP infusion was not recognized in IVF treatment [60, 61].
There were some limitations to this study. First, endometrial and blood samples were collected from the same women; therefore, given the limited number of extracted RNA samples, only microarray analysis and PCR array of undifferentiated and decidualized HESCs treated with and without PRP were performed, to identify detailed PRP-dependent genes of the PI3K-AKT signaling pathway. Second, the blood sample was kept in the transfusion pack in cold storage for 3–5 days. The anticoagulant, citrate phosphate dextrose adenine (CPDA), contributes to maintaining the function and viability of the platelets; however, according to a previous report [62], platelet count decreased by 20–30% after 3–5 days of storage in the CPDA-containing transfusion pack. Therefore, storage in a blood transfusion pack may affect the blood components and PRP. Third, the effect of PRP on endometrial epithelial cells was not analyzed in this study.
In summary, our observations suggest that intrauterine PRP infusion promotes tissue repair with transient inflammatory response, cell growth and migration, and antimicrobial effect in undifferentiated HESCs, whereas cell proliferation and the inflammatory immune response are attenuated during decidualization of the endometrium. Depending on the pre- and post-decidualization of the HESCs, PRP can regulate the reprogramming of inflammation and cell proliferation in an appropriate manner to support embryo implantation. However, PRP may have a suppressive effect on the decidual transformation of endometrium; thus, it may benefit only infertile women who have impaired intrauterine environment. Further investigation is required to elucidate the mechanism of action of PRP on the human endometrium.
{kind=link}