Immunobiotics feeding ameliorates MCD diet-induced NASH and LF in mice and is dependent on the gut permeability, gut microbiota, and AhR
The widely accepted methionine/choline-deficient (MCD) diet model that induces NASH and liver fibrogenesis in mice were used to determine the therapeutic potential of immunobiotics [31]. The mixture of three immunobiotic strains (live and heat-killed) was orally administrated from 0 day of MCD diet feeding (PL and PK) or after four weeks of MCD diet feeding (TL and TK) (Fig. 1A). The MCD diet-fed mice showed a significant decrease in body weight, liver and spleen weights, and an increase in the liver triglycerides (TG), serum GOT, and GPT levels. In contrast, the immunobiotic-fed mice showed significant improvement in their weights and a decrease in the liver TG and GPT (Fig. 1B). Mice fed the MCD diet developed micro and macrovascular steatosis and inflammation and increased NASH score and lipid accumulation in the liver (Fig. 1C and D). The liver sections from the mice administered immunobiotics (PL, PK, TL, and TK) had lower pathophysiological pictures of NASH than the MCD diet-fed mice. The PK group showed better results than the other groups. Feeding the MCD diet also led to significant increases in LF, which is characterized by increased collagen deposition, a-smooth muscle actin (a-SMA), and CD68 expressing KC and macrophages. (Fig. 2A and B). Collagen deposition and a-SMA are the staple markers for developing fibrosis and the activation of HSCs in the liver [3, 5]. The coadministration (PL or PK) immunobiotics had a significant effect on the reduction of MCD induced LF and the mRNA level of pro-fibrogenic markers, such as Col1A1, a-SMA, MMP-9, TGF-b, and TIMP-1 (Fig. 2A-C). The levels of these markers, except for MMP-9 in the TL and TK group, were significantly lower than in the MCD diet-fed group.
Immunoblotting analysis revealed the level of a-SMA to be higher in the liver samples of the MCD fed mice, but it was reduced by immunobiotics administration (Fig. 2D). Changes in the a-SMA protein levels were similar to the changes in the a-SMA mRNAs (Fig. 2C) and a-SMA immunostaining (Fig. 2A). Previous studies suggested the infiltration of hepatic T-helper (Th) cells during NAFLD and NASH development [8, 24]. Th17 cells with the specific IL-17 cytokine led to tissue inflammation and recruited leukocytes, which are reported to be higher in the NASH patients and NASH mice [11, 32]. Therefore, this study examined whether immunobiotics would modulate the frequencies of liver infiltrating Th17 cells and Treg cells in mice. The Th17 and T cells were measured by gating the singlets for CD45+ cells, followed by gating for CD4 cells. The frequencies of the IL-17-producing CD+T cells were higher in the liver tissues of the MCD-fed mice but lower in the immunobiotic fed groups (PL, PK, TL, or TK) mice (Fig 2 E). The mice fed the immunobiotics (PL or PK) showed a higher percentage of Treg, Tr1, and Th2 cells and a lower percentage of Th1 and CD3+CCR2+ cells than the MCD fed mice. This result suggests that the reduction of NASH-mediated liver fibrogenesis in the MCD diet-fed mice might be associated with the decreased infiltration of Th17 cells and increased infiltration of Treg cells.
In addition, to determine if the reduction of hepatic fibrogenesis in the immunobiotic-fed mice groups may cause by regulating the fibrogenic TGF-b signaling, the proteins associated with the TGF-b/Smad pathway in the liver tissues of mice were analyzed by western blot. Mice fed the MCD diet exhibited higher profibrogenic cytokine TGF-b levels than the chow-fed control group. Interestingly, immunobiotic administration (all groups) downregulated the TGF-b level in the liver tissues of mice to remarkably lower levels than in the MCD-fed mice groups (Fig. 2F). The phosphorylation of Smad3 was also down regulated in the immunobiotic-fed mice groups. A higher level of Smad7 was also observed in the liver tissues of the mice fed immunobiotics (Fig. 2F), suggesting that the immunobiotics beneficially modulated the TGF-b/Smad pathway to ameliorate the development of hepatic fibrogenesis in the NASH mice.
With regard to the inflammation-associated markers, the hepatic IL-17A, CCL5, TNF-a, IL-1b, CCL2, and TLR4 mRNA levels were higher in the MCD diet-fed mice than in the chow diet-fed mice groups (Fig. 3). The coadministration (PL or PK) immunobiotics significantly down regulated the expression of MCD-induced liver inflammatory markers and produced a higher level of anti-inflammatory cytokine IL-10 than MCD groups. In addition, MCD fed mice had higher liver mRNA levels of fibronectin, desmin, ICAM-1, MMP-2 and macrophage activation (YM-1), and fibroductular markers (K7, K19, and Sox9) (Fig. 4A and B). These changes were similar to the reports for mouse NASH livers [33]. Immunobiotic (PL, PK, TL, or TK) administration reduced the mRNA levels of ICAM-1 and Col1A2, and YM-1) and K19 than the MCD diet-fed mice (Fig. 4A and B), while there was no significant reduction found in the levels of desmin and Sox-9.
Immunobiotics alleviate the MCD diet-induced pathological changes in the intestine of mice
The liver tissue examination confirmed the protective role of immunobiotics, but it was unsure how the immunobiotics mediate their protective effects on hepatic fibrosis in mice. The loss of intestinal barrier integrity, increase in intestinal permeability, gut dysbiosis endotoxemia, and small intestinal bacterial overgrowth are standard features of NASH [34, 35]. Therefore, it was hypothesized that administering immunobiotics would improve the intestinal damage caused by the MCD diet in mice. At the end of MCD diet feeding, the length of the colon was shorter than chow diet-fed control mice (Fig. 5A). The mice fed the immunobiotics showed a significant increase in colonic length than the MCD diet-fed mice. The H&E histological examination revealed remarkable changes in the colonic structure of the MCD diet and immunobiotic-fed mice groups (Fig 6A). The colonic tissue of the control mice showed a normal morphological structure with intact mucosa and muscular layers. The mucosal layers were compact with clear crypt and goblets cells, and muscular layers were conserved without the infiltration of leukocytes (Fig 6A). The myenteric ganglia were full of neurons (Fig. 5B). The infiltration of leukocytes, eosinophils, no clear crypts, and loss of goblets and HuD positive neurons were observed in the colonic tissues of mice that fed the MCD diet. The colonic tissues of immunobiotic-fed mice showed a similar morphological pattern to that observed in the control mice. Collagen deposition and the mRNA level of Col1A1in the colonic tissues of the MCD-fed mice were lower than in control and immunobiotic-fed mice groups (Fig. 5C and D). The goblet cells density in the colon are important for maintaining the production of mucus and glycoproteins (mucins) that are dispersed widely throughout the mucosal epithelium and play roles in nutrient transport and epithelial cell protection [36]. Alcian blue (AB) and periodic acid-schiff (PAS) staining are widely used to stain mucins. Therefore, the acidic and neutral mucins were stained magenta in the colonic tissues of mice. AB-PAS straining showed that the MCD diet-fed mice had a low density of goblet cells with the depletion of mucus production compared to the control (Fig. 6B). The MCD diet mediated depletion was improved when the mice were fed the immunobiotic food. These results suggest that immunobiotic feeding improves the MCD diet-induced intestinal pathology in the mice.
Immunobiotics reduced the MCD diet-induced intestinal permeability and bacterial translocation in mice
Based on the colonic histology results, it was hypothesized that immunobiotics feeding would improve the intestinal barrier integrity in the MCD diet mice. An in vivo permeability assay was performed using FD4. The MCD diet increased the intestinal permeability, which was reduced by immunobiotic feeding (Fig. 6C). The intestinal barrier integrity is controlled by tight-junction proteins, mainly zonula occludens (ZO)-1, claudin-1, and occluding-1 [24]. The mRNA level of ZO-1, claudin-1, and occluding-1 were lower in the colonic tissue of the mice fed the MCD diet (Fig. 6C). As shown by the IHC and western blot study, a decrease in the level of these proteins was also observed in the colonic tissues of the MCD diet-fed mice compared to control and immunobiotic-fed mice (Fig. 6D-F). The PK group of mice had a higher ZO-1, claudin-1, and occluding-1 expression than the other immunobiotic groups (Fig. 6C). By contrast, the PL group of mice showed these proteins at a higher level than the other groups (Fig. 6E). The mice pre-treated with viable or heat-killed (HK) immunobiotic also increased the level of the TJ proteins before MCD diet feeding, but not higher than the PL and PK groups. Furthermore, this study investigated whether the MCD diet-induced intestinal permeability would increase the bacterial translocation to the liver tissue of the mice. The mice fed with the MCD diet exhibited gut bacterial translocation into the liver tissues (Table S1). The administration of immunobiotics impeded bacterial translocation. Only two of the five mice received viable immunobiotics (PL), and three of the five mice that received the other groups (PK, TL, TK) showed culturable bacteria. The chow diet-fed mice had no culturable bacteria in the liver tissue. These results suggest that using viable or heat-killed immunobiotics increased the TJ proteins to improve the intestinal barrier integrity and reduce the bacterial translocation in mice with MCD diet-induced LF.
Immunobiotics reduced serum endotoxin and Th17 levels and other inflammatory markers in mice
Previous studies reported that gut-derived endotoxin plays a critical role in inducing liver injury, particularly in the development and progression of NAFLD, NASH, and hepatic fibrogenesis [22, 37]. Gut-derived LPS translocate into the liver via portal circulation and activate TLR4 in the liver cells, resulting in the development of NASH and fibrosis [5]. The probiotic treatment reduced the serum endotoxin level and improved the gut-barrier integrity in the mice with choline-deficient/L-aminoacid-defined (CDAA) diet-induced NAFLD [37]. The mice fed the MCD diet had significantly higher serum endotoxin levels than the control mice (Fig. 6G). The immunobiotic (PL, PK, TL, or TK) treatment significantly reduced the endotoxin level in the serum of mice. There were no remarkable differences between the immunobiotics-fed mice groups. In addition, a significant increase in the serum and colonic mRNA levels of Th17 was observed in the MCD diet-fed mice compared to the control mice, but these increases were reduced significantly when the mice were fed the immunobiotics (Fig. 6G). In contrast, the immunobiotics treatment (PL or PK) elevated the serum and colonic mRNA expression levels of IL-10 in mice with MCD diet-induced LF (Fig. 6G). The other immunobiotics-fed group (TL or TK) mice also had a higher level, but not significantly, than the MCD-fed mice. Furthermore, the expression of other proinflammatory cytokines in the colonic tissues of mice was analyzed. The MCD diet increased the mRNA level of IL-6 significantly compared to the control mice (Fig. 6H). On the other hand, immunobiotic administration decreased the MCD diet-induced IL-6 expression in the colonic tissues of the mice but did not significantly down-regulate the mRNA level of TNF-a, IL-1b, and CCL2 compared to the MCD diet. The MCD diet upregulated the mRNA level of TLR4 significantly, but these were reduced by immunobiotics (Fig. 6H). Hence, the administration of immunobiotics reduced the serum and colonic inflammatory biomarkers by improving the intestinal barrier integrity or reducing intestinal permeability in mice with MCD diet-induced LF.
Immunobiotics modulated gut microbiota in mice with MCD diet-induced liver fibrosis
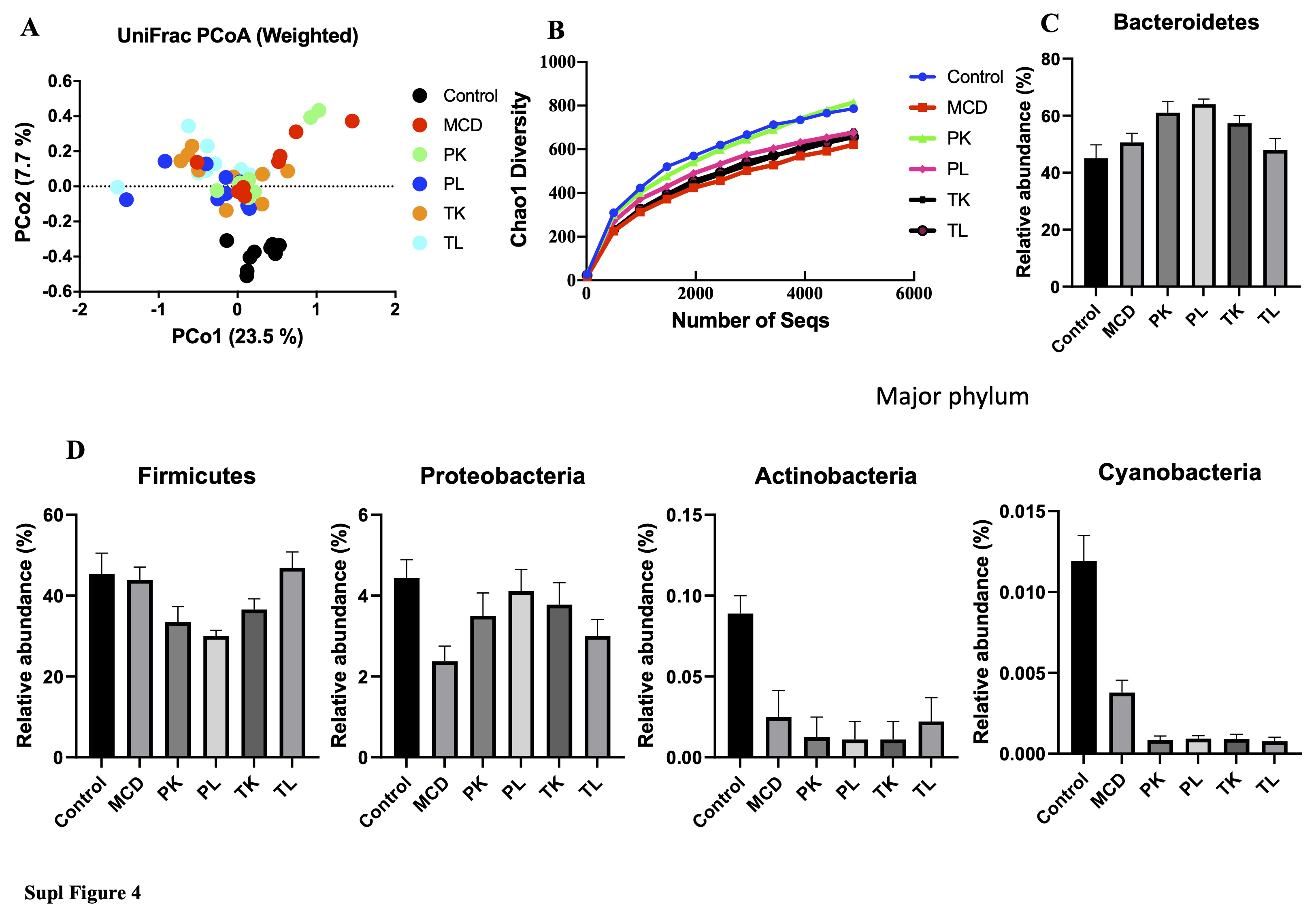
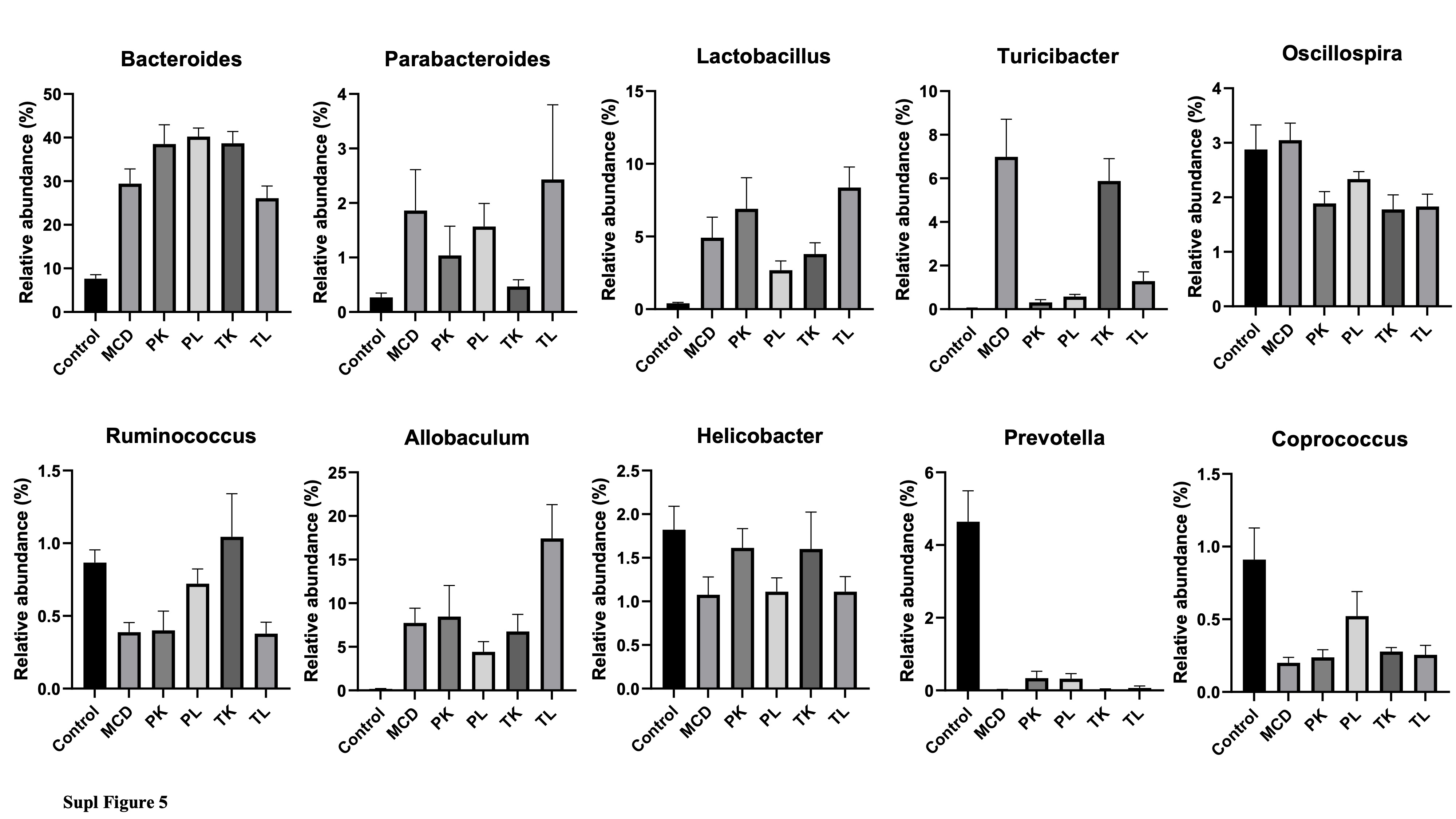
Several studies reported that the severity of fatty liver diseases is associated with the gut microbiota dysbiosis in humans and animals [38, 39]. Changes in the gut microbial composition were also observed in mice fed with a high-fat diet (HFD) and MCD diet [40, 41]. Therefore, it was hypothesized that the administration of immunobiotics would restore the changes in the MCD diet-induced gut microbiota in mice. The structural composition of the gut microbiota was determined by 16S rRNA sequencing of fecal samples from different groups of mice. A principal coordinate analysis (PCoA) based on the UniFrac unweighted and weighted showed that the structures of the microbial community segregated differentially among the groups (Fig. 7A and Fig 1SA). The MCD and immunobiotics fed groups were separated completely from the control group. The clusters of immunobiotic groups (PK and PL) were separated from the MCD groups, while the TK and TL groups formed clusters that were not completely distinct from the MCD group. These results suggest that the MCD diet induced a profound shift that could be restored using immunobiotic bacteria. The rarefaction curves reflected that the immunobiotic (PK and PL) fed groups were more diverse than the MCD fed group, while the TK and TL fed groups were not more diverse (Fig. 1SB). The MCD diet reduced the gut microbiota a-diversity metrics compared to the control and immunobiotic groups (PL and TL) (Fig 7C and D). The gut microbiota of all groups was classified as phyla, family, and genera that were different between the groups. The Bacteroidetes, Firmicutes, and Proteobacteria were the three dominant phyla observed in all groups. The relative abundance of the phyla, Bacteroidetes and Firmicutes, were similar in the MCD and chow diet-fed control groups (Fig 7D, Fig 1SC, and D), which is consistent with an earlier report [38]. On the other hand, Proteobacteria was lower in the MCD diet group than in the control group. Boursier et al. [39] reported similar results for patients with NASH and without NASH. Compared to the MCD group, an increase in the abundance of Bacteroidetes and Proteobacteria and a decrease in the abundance of Firmicutes and observed in immunobiotic fed groups (PK, PL, and TK). At the family level, MCD diet-fed mice had a higher abundance of Bacteroidaceae, Rikenellaceae, Lactobacillaceae, Turicibacteraceae, and Erysipelotrichaceae and a lower abundance of S24-7, Clostridiales, Lachnospiraceae, Ruminococcaceae, and Prevotellaceae compared to the control group (Fig. 7E). Similarly, the abundance of fibrosis-associated Ruminococcaceae was decreased when fibrosis could become more severe in non-obese individuals42. Nevertheless, immunobiotic administration altered the pattern of the bacterial abundance at the family level compared to MCD groups. At the genus level, compared to the control group, the relative abundance of Bacteroides, Parabacteroides, Lactobacillus, Allobaculum, and Turicibacter were higher, whereas Ruminococcus, Coprococcus, and Prevotella were significantly lower in the MCD groups (Fig. 3SA and B). Similar results were observed in the non-obese subjects with severe fibrosis. The administration of Ruminococcus alleviates liver damage in NAFLD mice fed with the MCD diet [42]. On the other hand, immunobiotic groups (PL and PK) had a different pattern of gut microbiota at the genus level than the MCD group, suggesting that immunobiotic feeding beneficially alters the gut microbiota to protect against fibrosis.
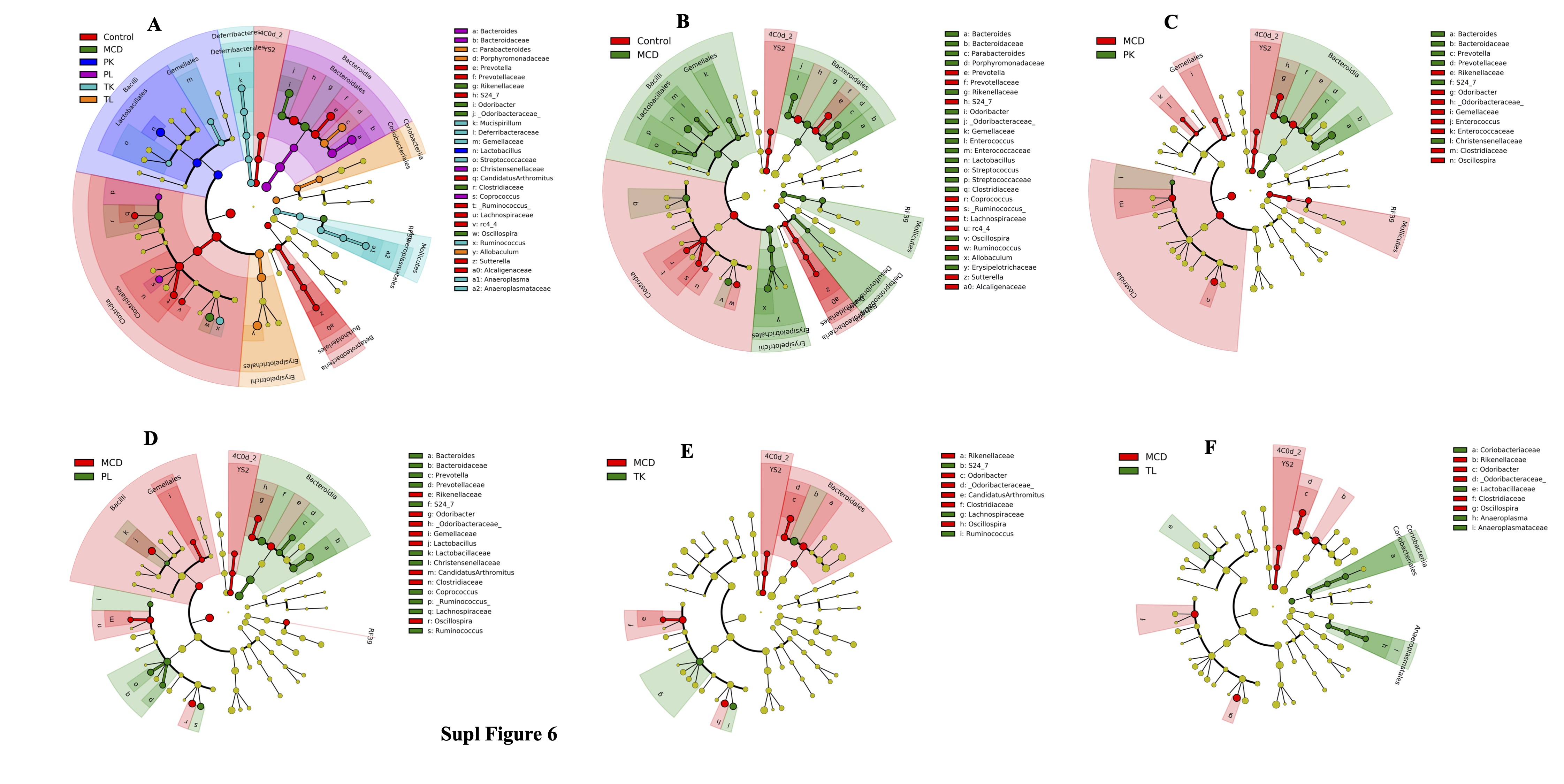
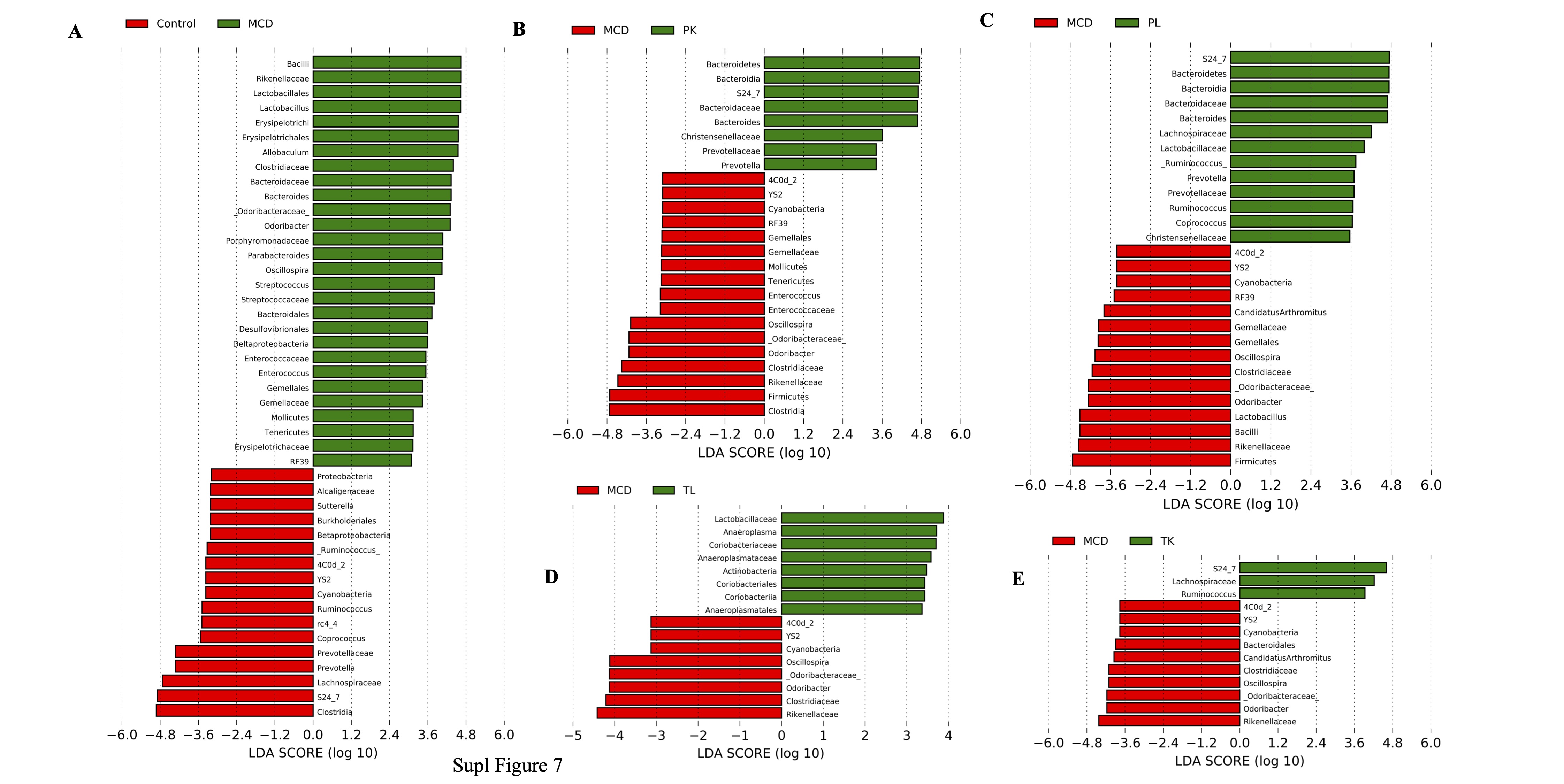
In addition, specific taxa that respond to MCD diet and immunobiotic administration were examined using the LDA effect size (LEfSe) assay to determine the differences in the microbial abundance among the groups and each other. The results showed that 55 microbial taxa at the genus level were identified as markers that discriminate the gut microbiota among the groups (Fig. 7F, Fig. 3SA). In the MCD group, the microbial taxa changes after the MCD diet were dramatic compared to the chow diet-fed control group. Interestingly, immunobiotic-feeding groups had different genus levels of microbial taxa compared to the MCD group. In particular, Lactobacillales and Lactobacillus, which belong to the family Lactobacillaceae and phylum Firmicutes, may play a role in alleviating the development of fibrosis, were higher in the PK group compared to the MCD group. In contrast, Bacteroides in phylum Bacteroidetes were enriched in the PL group, while the abundance of Erysipelotichia (a producer of short-chain fatty acid) in phylum Firmicutes was higher in the TL group compared to the MCD-diet group. The number of microbial taxa was different when compared with each other (Fig. 3SB-F, Fig. 4SA-E).
This study examined the correlation of the gut microbiota with the disease markers to study the impact of the gut microbiota on disease. The abundance of Lactobacillus and Allobaculum (higher in PK and TL groups) was positively associated with body weight and liver and spleen weight (Fig. 7G). In addition, a genus Bacteroides enriched in the immunobiotic groups had a positive association with the body weight and liver weight. Moreover, the abundance of Odoribacter was positively correlated with the body weight, liver, and spleen weight and negatively correlated with the liver TG, GOT, GPT, and serum endotoxin. By contrast, Prevotella (lower in the MCD diet-fed group) and Suttrella were negatively correlated with the body weight, liver, and spleen weight, while positively correlated with the GOT, GPT, and serum endotoxin levels. Coprococcus and Ruminococcus were lower in the MCD diet-fed group. They had a moderate negative correlation with body weight, liver, and spleen weight and a moderate positive correlation with the serum endotoxin and GPT levels (Fig. 7G). The genus Oscillospira had a strong negative correlation with the liver TG, GOT, GPT, and serum endotoxin levels. Previous studies showed that the MCD diet lowered the body, liver, and spleen weights, and increased the liver TG, GOT, GPT, and serum endotoxin levels, which are major risk factors for liver injury by inducing inflammation in the host [37, 43]. A strong correlation was observed between these factors and the gut microbiota, which may participate in the development of LF.
Immunobiotics activated AhR in the colonic and liver tissues of mice and in vitro cells
Overall, immunobiotics modulate the MCD diet-mediated gut microbiota in mice, but it is unclear how the modulation of gut microbiota by immunobiotics would alleviate LF. Studies reported that the expression of AhR, production of AhR ligands, and AhR pathway activation are reduced in the gut of patients with intestinal diseases [26]. Modulation of the gut microbiota by Lactobacillus increased the production of AhR ligands and AhR activation, which alleviated the intestinal pathology in mice [26]. The production of sufficient AhR ligand is critical for restoring the composition and function of the gut microbiota [44]. AhR signaling protected against intestinal diseases and prevented HSC activation and liver fibrosis in mice [21]. Therefore, this study next examined whether immunobiotics feeding increases the expression of AhR and AhR signaling activation in both intestinal and liver tissues of mice. Interestingly, the protein and mRNA levels of AhR and its signaling proteins (Cyp1b1 and Cyp1a1) were higher in the colonic tissue of mice fed the immunobiotics than in the MCD diet alone (Fig. 8A and B). A similar pattern of AhR and other proteins was found in the AhR colonic cells treated with fecal extracts derived from mice fed the immunobiotics (Fig. 8C). AhR activity was found to be increased in HT-29 cells treated with feces from PL and PK group of mice (Fig. 8C). Furthermore, the mRNA levels of these proteins were higher in the liver tissue of the immunobiotic-fed mice groups (Fig. 8D). On the other hand, AhR at the protein level was higher in all immunobiotic groups, while Cyp1a1 and Cyp1b1 were lower in the PL group compared to the MCD group (Fig. 8E). An in vitro study also showed a similar pattern of AhR protein in LX-2 cells incubated with a fecal extract from the immunobiotic-fed mice groups (Fig. 8G). An immunohistochemistry study showed that the liver tissues of mice administered with immunobiotics had higher AhR positive cells than the MCD group (Fig. 8F). These results suggest that immunobiotics positively modulated gut microbiota to produce a higher level of AhR ligands and AhR pathway activation, which may play a crucial role in reducing LF in the MCD diet-fed mice.
{kind=link}
{kind=link}
{kind=link}
{kind=link}