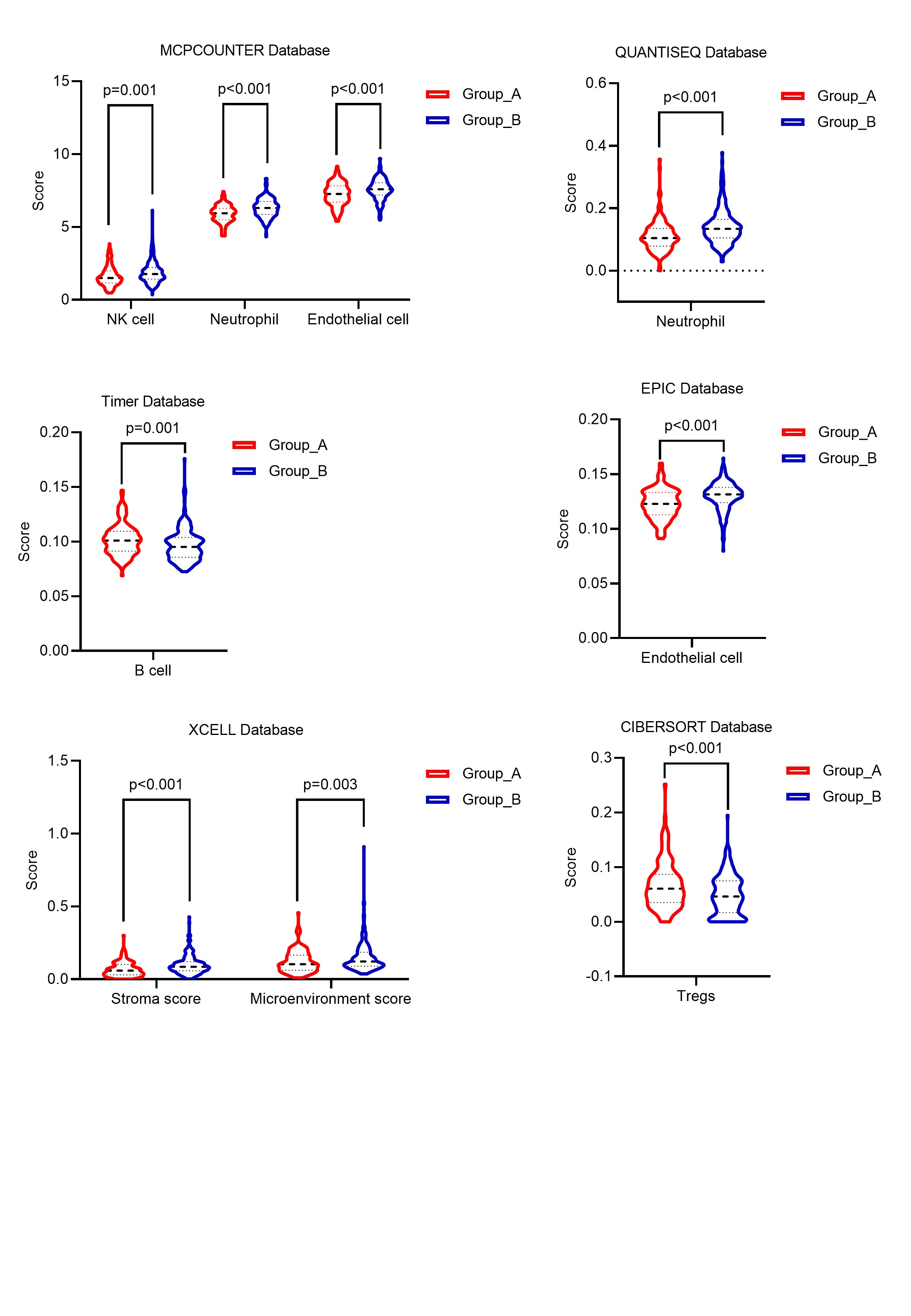
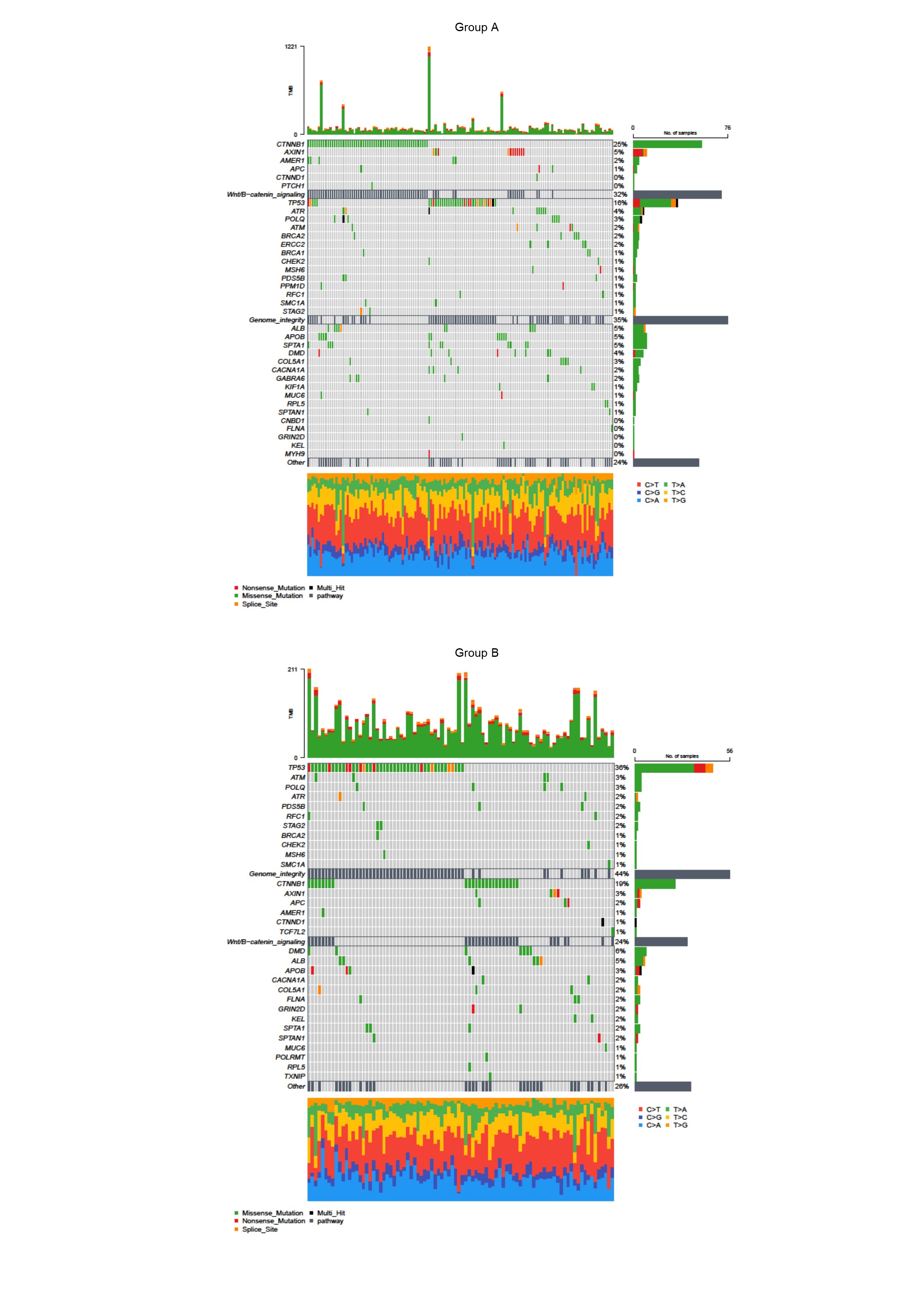
It is well-established that aberrant expression of core PcG subunits contributes to tumor initiation and progression. In HCC, overexpression of EZH2 represses miR-622 through H3K27me3 deposition and results in CXCR4 upregulation and unfavorable prognosis, while BMI1 enhances TGFβ2/SMAD pathway and facilitates tumor cell proliferation and cell cycle progression [25, 26]. In recent years, accumulating evidence suggested that other non-core accessory proteins alternatively constituting the PcG complex can also facilitate the pro-tumor process. For example, euchromatic histone lysine methyltransferase 2 (EHMT2), an alternative subunit of PRC1.6.has been recently reported to facilitate HCC progression and aggressive features through epigenetic silencing of tumor suppressor genes [27]. In addition, although PR-DUB and PhoRC are scarcely reported in the field of HCC study, based on their close relationship with PRC1/2, we supposed that they are also functional in the epigenetic classification of HCC [28]. Therefore, in this study, aiming to analyze the subtype of HCC from the perspective of epigenetic regulation of the PcG complex in a more comprehensive picture, we included all the core and non-core proteins of four PcG complexes. Patient in two subgroups has a significantly distinct prognosis and exhibit different biological features, which is related to aberrant PcG complex activity and epigenetic landscape.
Abnormal histone modifications, especially methylation and ubiquitination, are associated with poor prognosis of cancer patients and decides biological phenotypes of tumor tissue [29–31]. It has been reported that histone methylation is associated with various malignant features like vascular invasion, large tumor size, multiplicity as well as relapse, and a previous research has also proved the tumor-promoting role of histone ubiquitination in HCC [32, 33]. For HCC, aberrant expression of PcG proteins like BMI1, CBX8, and EZH2, is mainly responsible for the deposition and maintenance of pro-tumor histone modification [26, 34]. In our study, we identified two subtypes of HCC by the analysis of the expression pattern of PcG-related genes, and CBX8 and EZH2 are upregulation in group A. Thus, we referred that Group A HCC had elevated histone methylation and ubiquitination. According to survival analysis, patients in group A have a worse prognosis, especially in young and stage Ⅲ/Ⅳ subpopulation. These results suggest that the poor prognosis of Group A patients is related to pro-tumor histone modification which is consistent with previous studies.
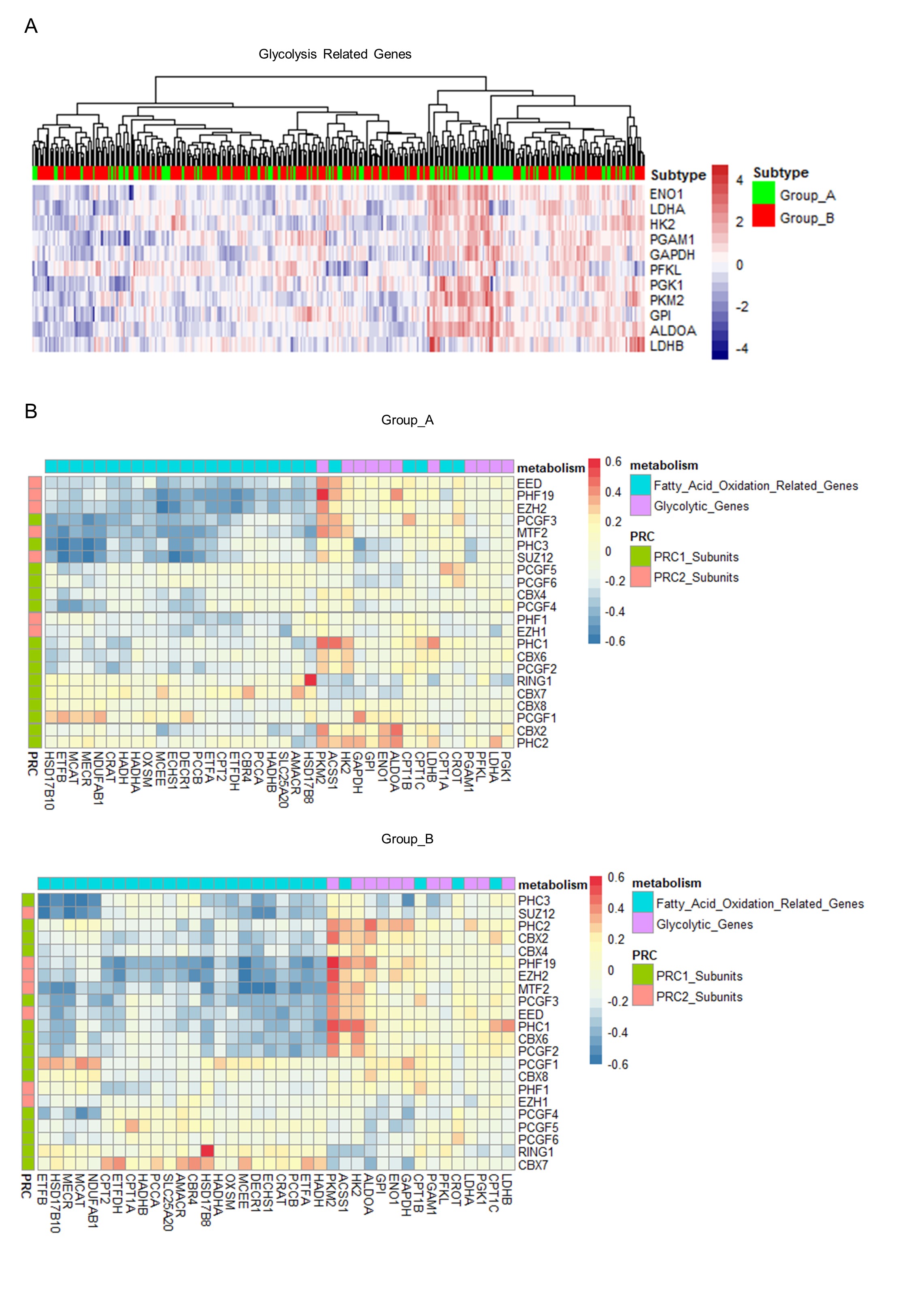
According to our informatics analysis, HCC cells in two subgroups have distinct characteristics in many aspects, especially cell metabolism. We found that patients in Group A have lower expression of the fatty acid metabolism gene, and the downregulation of these genes can be abolished by the PRC1/2 inhibitors. Consistently, the capacity of fatty oxidation and accumulation of HCC cells were also promoted after PRC1/2 inhibitors treatment. In addition, our results showed the increased dependence on lipid and sensitivity to glucose deprivation of PRC1/2- inhibited HCC cells. In recent years, many studies have suggested that metabolic homeostasis is crucial for HCC cells in a setting of reduced nutrient availability, which can be affected by the epigenetic regulation [35–37]. Recently, Li et al proved that the deficiency of SET domain-containing 2 (SETD2), a histone methyltransferase, could cause downregulation of H3K36me3 and cholesterol efflux genes, which led to lipid accumulation and HCC development [35]. In our study, we focused on H3K27me3 and H2AK119ub which are regulated by PcG complexes, and found these histone modifications have a broad influence on the aspect of fatty acid metabolism. Intriguingly, downregulation of H3K27me3 and H2AK119ub alters HCC cells` preference of energy source from glucose towards fatty acid and make them more fragile under the shortage of glucose, which impairs their capacity of metabolic adaption. However, whether epigenetic regulation of metabolic adaption could affect the initiation and progression of HCC needs further investigation.
The drugs targeting mutant PcG proteins such as EZH2 have been extensively investigated, and several of them have been applied to preclinical research of hematological malignancy treatment [38, 39]. However, in solid tumors, aberrant function and excessive enzymatic activity of PcG proteins are usually derived from their overexpression instead of their mutation, where the efficiency of PcG targeting drugs is limited. To solve this problem, PcG protein targeting drugs can be combined with immune therapy, conventional chemotherapy, or other targeting therapy [40, 41]. Here, we combined oxaliplatin, a widely used chemotherapeutic drug for gastrointestinal malignancy treatment, with PRC1/2 inhibitors respectively or synergistically. Our data showed that the efficiency of combined therapy to induce HCC cell apoptosis in vitro and inhibit tumor progression in vivo. In addition, our study proved that synergistic inhibition of PRC1 and PRC2 can bring higher sensitivity to oxaliplatin, and similar therapeutic strategy have recently being reported in glioblastomas treatment where the investigators successfully used molecular inhibitors of BMI1 and EZH2 to control proneural and mesenchymal tumors under the limits of detection [42]. Targeting PcG proteins dissolves the pro-tumor histone modification and enhances the sensitivity of cancer cell to other treatments.
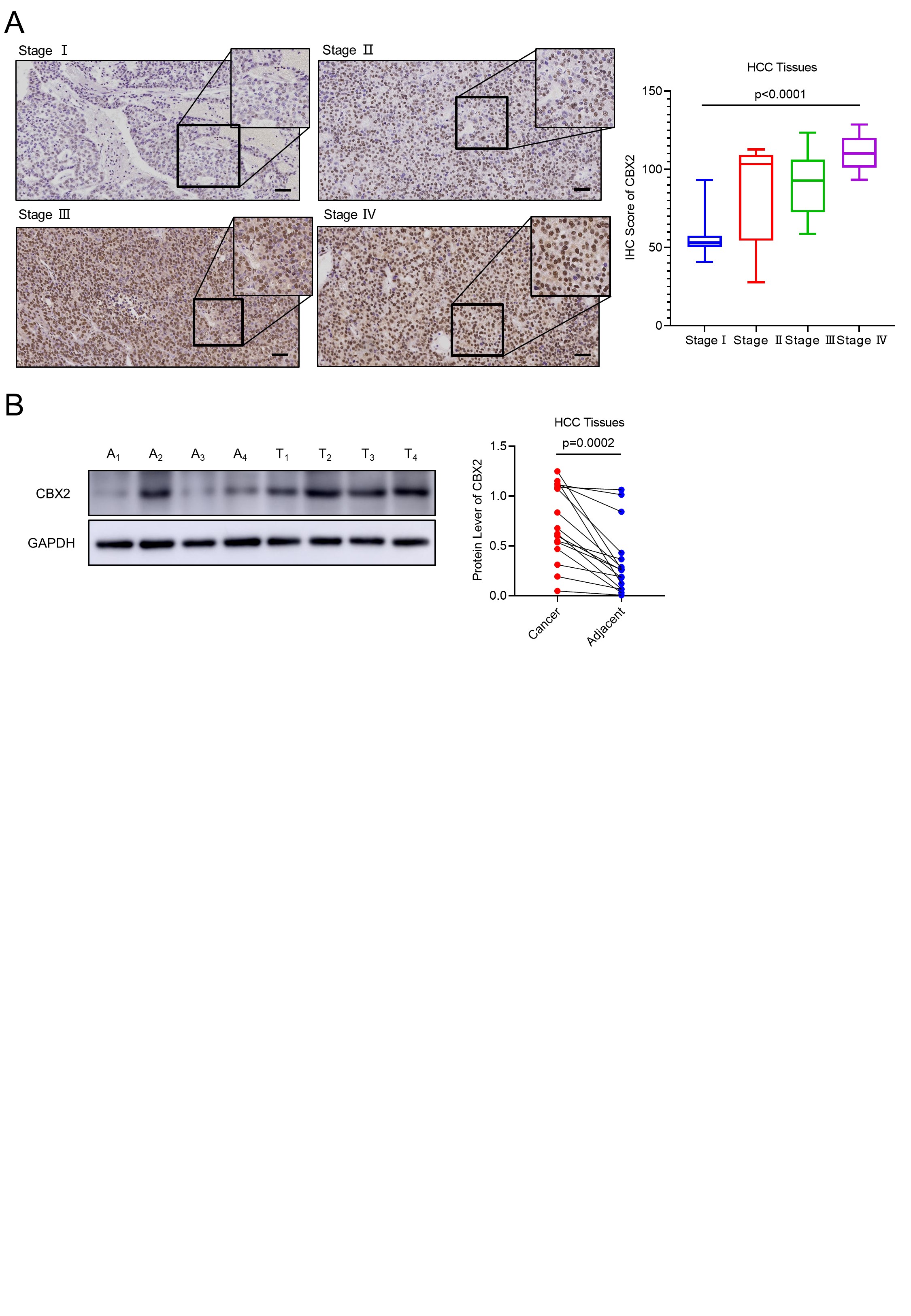
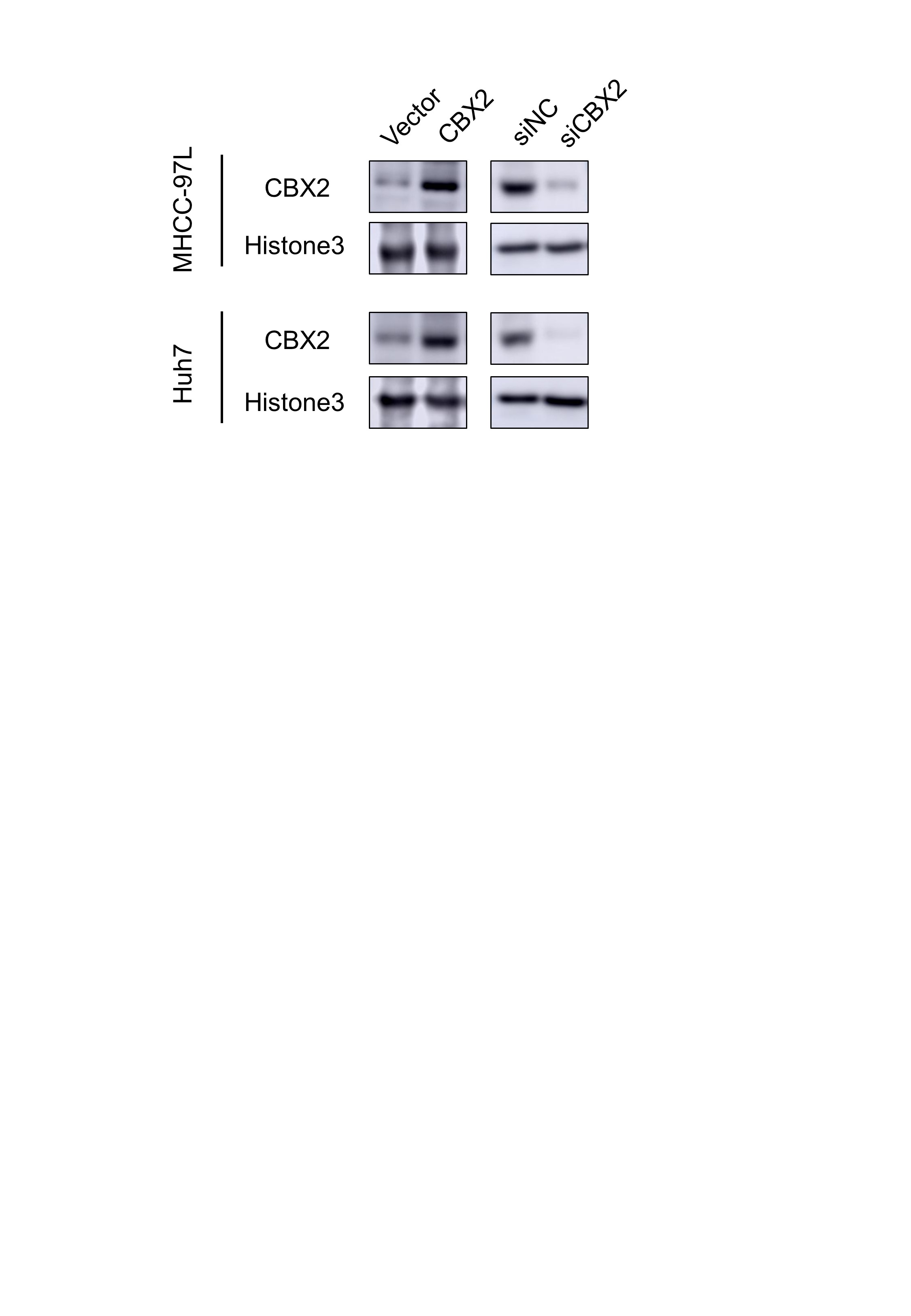
In our study, we proved that overexpression of CBX2, the epigenetic reader, contributes to overexpression of histone methylation and ubiquitination by promoting the activity of PRC1/2 in HCC cells. As a “bridge” of PRC1 and PRC2 intercommunication, CBX2 recognizes the H3K27me3 marker and recruits other subunits of PRC1 facilitating histone modification of H2AK119ub which can be recognized by PRC2, which induces chromatin compaction for deep silence of genes [43, 44]. A previous study has reported the extranuclear function of CBX2 to activate the YAP pathway in HCC progression [19]. Here, our result showed the intranuclear function of CBX2 to promote the H3K27me3 and H2AK119ub deposition. A recent review summarized the therapeutic potential of targeting CBX2 and its advantages beyond directly targeting EZH2 [24]. Consistently, our study provided evidence that knockdown of CBX2 could simultaneously alleviate pro-cancer histone modification, and this therapeutic strategy may especially benefit HCC patients who bear an aberrant epigenetic landscape like subtype Group A patients.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}