FIR contains the highly conserved acidic string region that potentially interacts with P62 of TFIIH.
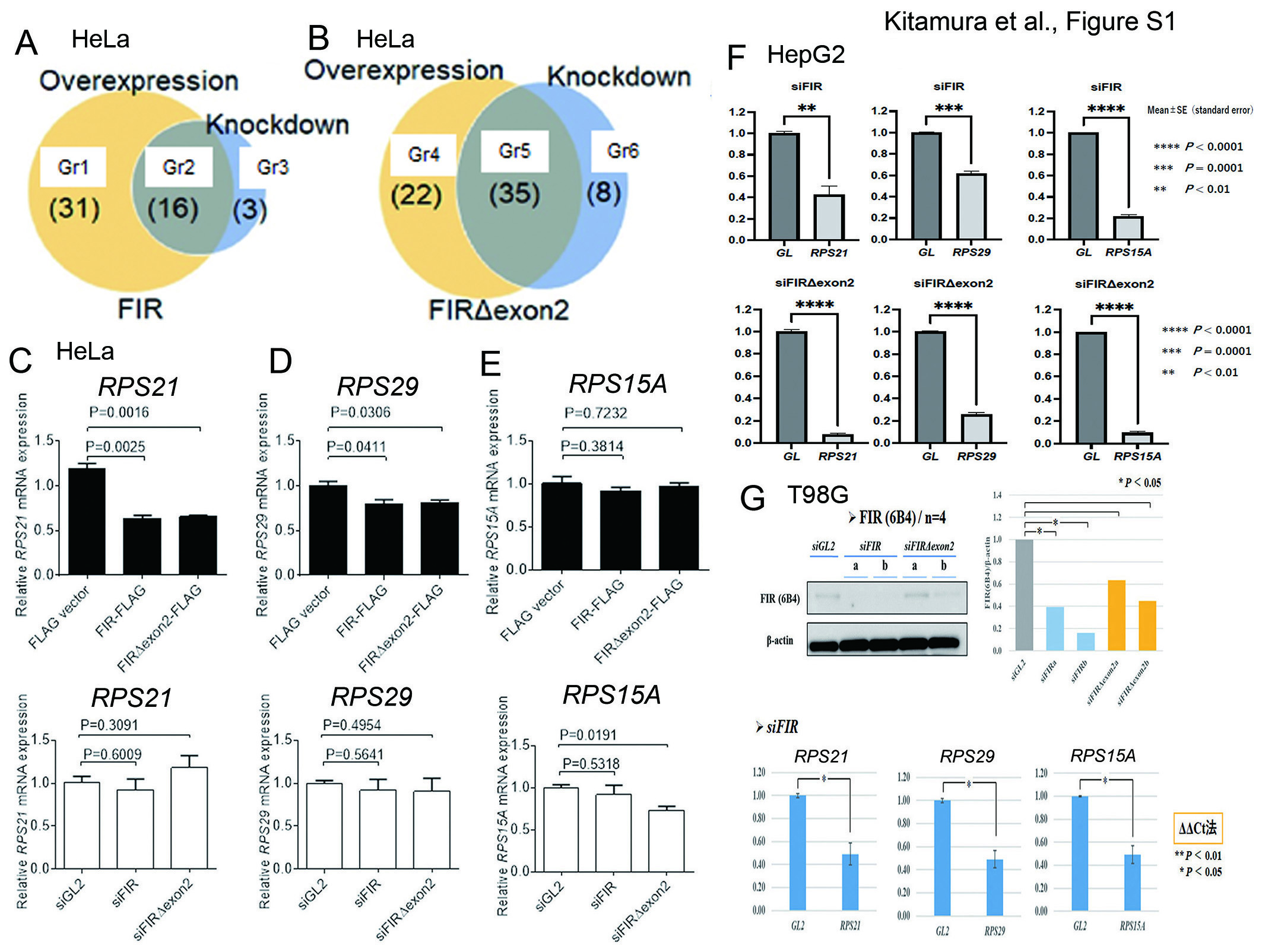
The N-terminal tail (NTT) of RPB6 interacts with the PH (pleckstrin homology)domain of the P62 subunit of TFIIH8. These FIR family members (FIR, PUF60, and FIRΔexon2) contain three RRMs (RNA recognition motifs; RRM1, RRM2, and RRM3) (Figure 1A,1B). RRM1 binds to the c-myc promotor and RRM2 binds to a transcriptional activator4,16,17. FIR contains the highly conserved acidic string region (376-KKEKEEEELFPESERPEM-394) between RRM2 and RRM3/UHM (Figure 1A). As the crystal structure is unavailable for the acidic string region, this region is flexible and easily changes its conformation (Figure 1B). Partial domain structures of FIR, RRM1 (PDB#:2QFJ)18, RRM2 (PDB#:2KXH)16, and UHM (PDB#:3DXB)19, were previously elucidated by X-ray crystallography. The FIR protein binds ssDNA of FUSE as a dimer in which only the RRM1 and RRM2 domain of each subunit interacts with ssDNA (Figure 1C)18. RNAPI, II, and III contain the common NTT in RPB6 (Figure 1D). Interestingly, these acidic strings are highly conserved among FIR, RPB6, UVSSA, XPC, TFIIEα, TP53, and DP1 (Figure 1D). FIRΔexon2, a splicing variant form of FIR that lacks exon2, is overexpressed in cancer cells as a dominant-negative form of FIR5,6,20,21. Given, FIR is crucial for RNAPI expression in cellular development, FIR affects RNAPI expression in a variety of differentiated cells. For this purpose, HepG2 cells (human hepatoblastoma cells), 98G cells (human glioblastoma cells), HCT116 cells (human colorectal cancer cells), and Hela cells (human cervical squamous carcinoma cells) were examined in this study. As anticipated, the knockdown of FIR expression by siRNA led to significant suppression of P62 in HepG2 cells (Figure 2A) and in T98G cells (Figure 2B) as well as in HCT116 cells (Figure 2C). The significant suppression of P62 was determined by densitometry analysis, with β-actin used as an internal control (Figure 2D). These results indicated that FIR is required as a scaffold protein for the stable expression of P62 and the E (glutamic acid)-rich sequence (Figure 1D). Considering that the E-rich acidic amino acid sequence along with the aromatic (F) and hydrophobic (L) amino acids of FIR interacts with P62 of TFIIH (Figure 1D), the expression of RNAPI and rRNA should be affected by FIR and FIRΔexon2. In the case of FIR, 31 (Gr 1) + 16 (Gr 2) = 47 genes were affected by the overexpression of FIR-FLAG, whereas 16 (Gr 2) + 3 (Gr 3) = 19 genes were affected by the knockdown of FIR by siRNA in HeLa cells (Table 1). Similarly, in the case of FIRΔexon2, 22 (Gr 4) + 35 (Gr 5) = 57 genes were affected by the overexpression of FIRΔexon2-FLAG, whereas35 (Gr 5) + 8 (Gr 6) = 43 genes were affected by the knockdown of FIRΔexon2 by siRNA (Table1).
Evaluation of the interaction between FIR and P62 of TFIIH by the isothermal titration calorimetry thermogram
The isothermal titration calorimetry (ITC) thermogram, showing the titration curve of P62 of TFIIH with FIR, suggested a molecular interaction between the two proteins. The exothermic peaks appeared with the earlier injections, and the exothermicity of the injection decreased with subsequent injections. The dissociation constant was calculated as Ka = 2.34 × 107 M−1 from the ITC thermogram (Figure 3A). The binding affinity energy between P62 of TFIIH and FIR was enthalpically favorable as the ITC thermogram revealed that the interaction was exothermic. However, the binding of P62 of TFIIH and FIRΔexon2 was not significant (Figure 3B). These results indicated that FIR interacts with P62 of TFIIH, and the interaction capability with P62 is potentially different between FIR and FIRΔexon2. Why do both FIR and FIRΔexon2 display significant effects on rRNA expression, although only FIR interacts with P62 expression? On possibility is that FIR forms heterodimer with FIRΔexon2 upon RRM1 and RRM222; therefore, FIRΔexon2 potentially affects P62 expression through forming heterodimer with FIR on RRM1 and RRM2.
FIR and FIRΔexon2 dynamically stimulated rRNA expression
The expressions of RPL30, RPL37A, RPL38, RPS14, and RPS29 mRNAs were decreased significantly by the overexpression of both FIR-FLAG and FIRΔexon2-FLAG. Similarly, the rRNA expressions of RPL19, RPL37, RPS6, RPS10, RPS15A, and RPS21 were decreased significantly by the overexpression of FIRΔexon2-FLAG in HCT116 cells (Figure 4A). The knockdown or overexpression of FIRΔexon2 significantly decreased the rRNA expressions of RPL19, RPS6, and RPS10 by siRNA. (Figure 4B), indicating FIRΔexon2 is required for sustained rRNA expressions of RPL19, RPS6, and RPS10. The rRNA expressions of RPL37, RPS10, RPS15A, RPL6, RPL29, and RPL23A were increased by the knockdown of both of FIR and FIRΔexon2. Furthermore, we performed RT-PCR to analyze the expression of RPS15A, RPS21, and RPS29 in HeLa cells (Figure S1C, D, and E). The relative RPS21 and RPS29 mRNA expressions were decreased significantly with the overexpression of both FIR and FIRΔexon2-FLAG (top, Figure S1C, D, and E). In contrast, the relative RPS15A mRNA expression was decreased significantly by siFIRΔexon2 (bottom, Figure S1C, D, and E). Knockdown of FIR or FIRΔexon2 by siRNA affected the mRNA expressions of RPS15A, RPS21, and RPS29 in HepG2 cells (Figure S1F). Knockdown of FIR by siRNA affected the mRNA expressions of RPS15A, RPS21, and RPS29 in T98G cells (human neuroblastoma cells) (Figure S1G). The results of comprehensive RNA-seq analysis demonstrated that numerous rRNA-processing proteins were prominently affected by FIR or FIRΔexon2 (Table S1). Numerous RPs were coimmunoprecipitated with FIR and FIRΔexon2 (Table S2)22, 23, suggesting that FIR and FIRΔexon2 coexist with RPs and participate in dynamic rRNA expression.
FIR or FIRΔexon2 affected the dynamic expression of various genes for rRNA synthesis.
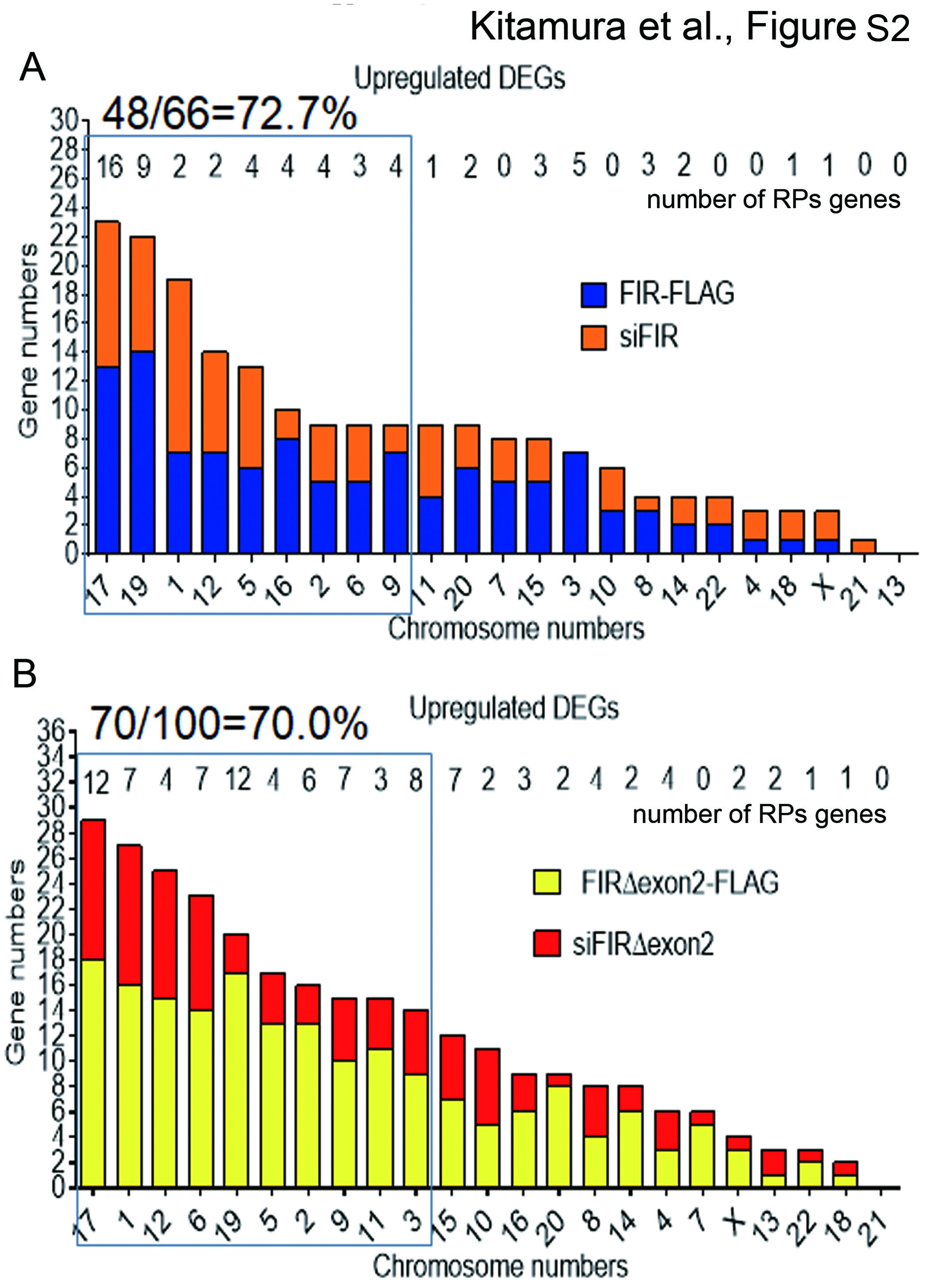
The RPs that constitute the ribosome are generated from the rRNAs transcribed from rDNA by RNAPI in the nucleolus. Given that FIR participates in dynamic rRNA transcription, the accessibility of FIR to genes for rRNA synthesis on the specific chromosome would be related to RP synthesis. Overlapped genes affected by both overexpression or knockdown of FIR (Figure S2A) and FIRΔexon2 (Figure S2B) were located on chromosomes 17, 12, 19, 5, 2, 6, 9 and 1 (Figure S2). On the other hand, five human acrocentric chromosomes (13-15, 21 and 22) contain tandemly repeated clusters of genes for rRNA in the satellite region of their short arm24. Robertsonian translocations, centric fusion of the two chromosomes among above five acrocentric chromosomes, indicated 20% lower number of active ribosomal genes than those in the controls24. In such cases, insufficient rRNA synthesis resulting in impaired RiBi called “ribosomopathies”2. These results strongly suggested that FIR and FIRΔexon2 targets genes for rRNA synthesis on specific chromosomes that causes “ribosomopathies”.
FIR or FIRΔexon2 in affects the genes responsible for rRNA expression at least partially through a c-Myc-independent mechanism.
c-Myc enhances RNAPI activation by binding the rDNA promoter25. c-Myc regulates protein synthesis by stimulating RPs; RPS and RPL, small and large ribosomal subunits, respectively26. As FIRΔexon2 suppressed c-myc gene transcription as a dominant-negative form of FIR5, c-Myc expression was upregulated by the knockdown of FIR by siRNA. The affected genes for rRNA due to both increased and decreased expression of FIR were RPL26 and RPS27, and RPL39. The affected gene transcription due to both increased and decreased expression of FIRΔexon2 were RPL11, RPL3, RPL41, RPL7, RPL7A, RPS25, and RPS3A (Figure 5A, 5B). However, the majority of RPS and PRL genes were independent of c-Myc expression (Figure 5C, D). Here, c-Myc expression was evaluated using the average of FPKM in RPL or RPS genes that were commonly detected in duplicated experiments by the knockdown of FIR or FIRΔexon2 siRNA in HeLa cells under a cut-off level FDR of <0.05 (Table S2). These results indicated that FIR or FIRΔexon2 engages in rRNA expression at least partially through a c-Myc-independent mechanism (Figure 5).
Cell growth inhibition by an in silico screened chemical BK697 against FIR and FIRΔexon2
Small molecular weight chemical compounds potentially bound to FIR and or FIRΔexon2 was previously identified from the Natural Product Depository (NPDepo) at RIKEN (Saitama, Japan), which was a collection of the isolates from natural products (Table S3) 13-15. A small molecular weight chemical that potentially interacting with FIR was identified by NPDepo screening (A01 of Table S3). From computer screening to search synthesized chemicals that mimicking the structure of the identified natural chemical compound A01 (1,4a-Dimethyl-2,3,4,4a,9,9a-hexahydro-1H-fluorene-1,9-dicarboxylic acid) (Table S3) by Namiki database (Namiki Shoji Co., Ltd., Tokyo, Japan) (Figure 6A). The commercially available chemicals were screened that composed of the structure of A01 (Figure 6B-(A)) and identified some chemicals (Figure 6B-(B), (B’)). Chemical skeleton of the two synthesized compounds (Figure 6B-(B), (B’)) were regarded as a Trp (W) and Asp (D) combination (WD motif) mimicking form (Figure 6B-(C))27. Affiliated chemicals were screened and indicated that BK697 (Figure 6B-(D)) effectively suppressed HepG2 cell growth with IC50 of 17.8 mM (Figure 6C). BK697 suppressed FIR and FIRΔexon2 expression in HepG2 cells of their protein (Figure 6D) and mRNA level (Figure 6E). c-Myc expression was decreased along with FIR and FIRΔexon2 suppression by BK697 in Hela cells (Figure 6F). RPS6, RPS10, RPS15A, RPL30, and RPL23A expression was reduced by BK697 with IC50 of 17.8 mM in HepG2 cells (Figure 6G). Clinically, BK697 and its derivatives are potential candidate anticancer drugs for cancers targeting FIR and FIRΔexon2 suppression.
FIR and FIRΔexon2 potentially interact with RPs and monitor their expression
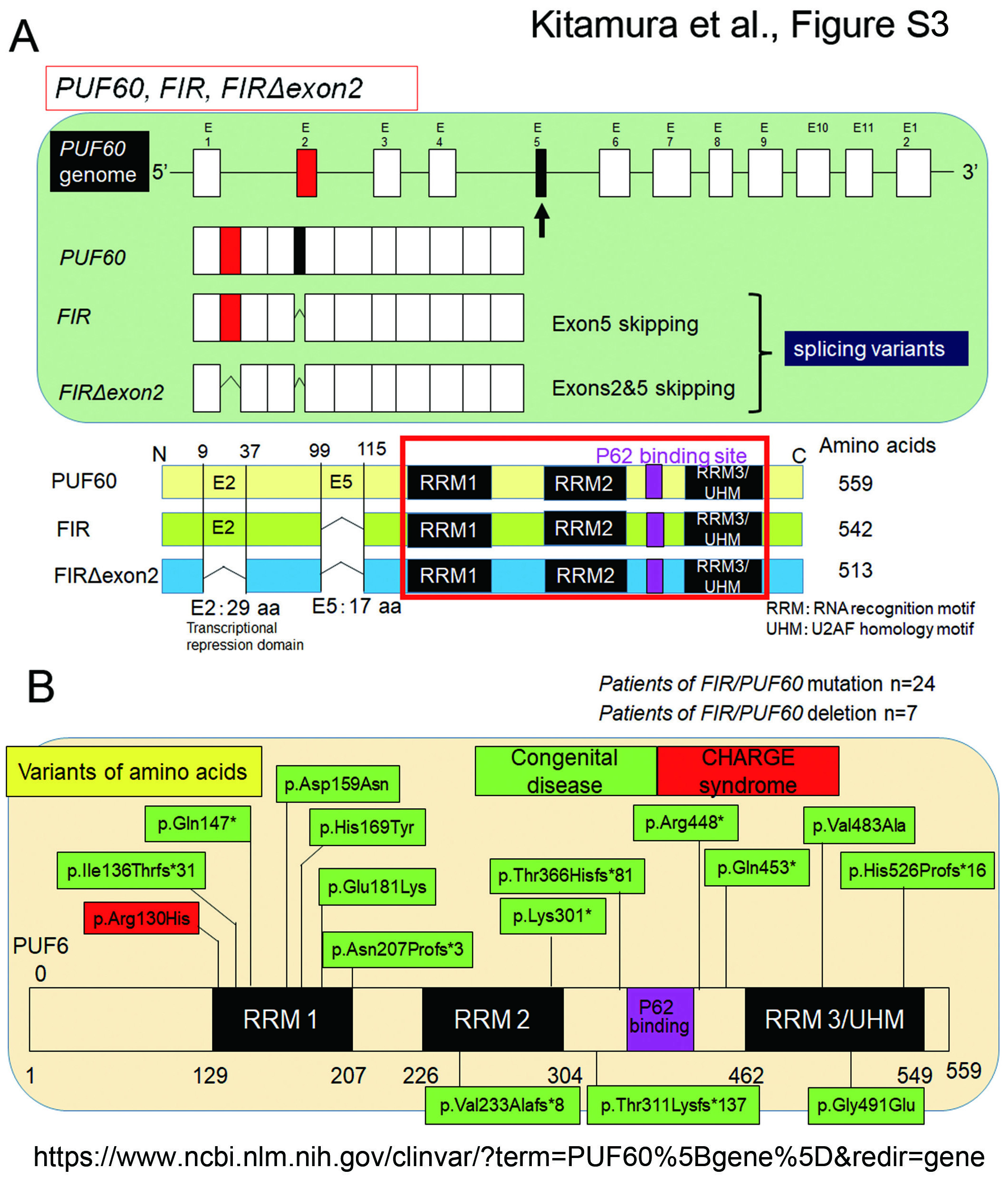
RPs have been identified as coimmunoprecipitated proteins with FIR and FIRΔexon2 (Table S1) 20,22,23. This supports the notion that FIR and FIRΔexon2 stimulate rRNA expression through P62 binding site between RRM2 and RRM3/UHM (U2AF homology motif) (Figure S3A) and potentially interact with RPs (Tables S1 & S2). Remarkably, the pathogenic germline variants of FIR/PUF60 gene have been accumulated as a cause of Verheij syndrome or CHARGE syndrome, which are congenital diseases related to neural development (Figure S3B). Since ribosome synthesis is critical for neural development28, FIR family potentially bridges neural disease and cancer development through dynamic rRNA stimulation (Figure 7).
FIR and FIRΔexon2 dynamically stimulate RPs and cellular proliferative gene expression.
FIR/PUF60 and c-Myc are located at 8q24.3. FIR contains the highly conserved acidic amino acid string region, a similar sequence of the RPB6 subunit that interacts with P62 of TFIIH. FIRΔexon2, unlike FIR, interacts with P62 of TFIIH and stimulates RP small subunits (RPSs) and those of large subunits (RPLs) (Figure 7) in the nucleolus. Previous studies reports have supported that the FIR family potentially interacts with RPs20,22,23. Therefore, the FIR family is a bona fide modulator of rRNA transcription of RP via P62/TFIIH. Consequently, FIR and FIRΔexon2 monitor the expression of RPs and participate in RiBi by regulating rRNA expression through the P62 PH domain of TFIIH. These results suggested that FIR and FIRΔexon2 involve in the rRNA expression and cellular proliferative gene expression.
{kind=link}
{kind=link}
{kind=link}