Identification of OsMGT genes and their characteristics
In the current study, 23 non-redundant putative MGT proteins, including 11 NIPA, 9 MRS, and 3 CorA members, were recognized in the genome of rice (Table 1). The predicted physicochemical characterizes in MGT proteins revealed high diversity. For example, the molecular weight (MW) ranged from 8.50 (in Os11g0197400) to 62.91 kDa (in Os01g0664100). Most of the identified MGT proteins in rice had acidic nature, because of the isoelectric point (pI) values lower than 7, and pI values ranged from 4.61 (in Os04g0430900) to 11.43 (in Os10g0545000). The predicted instability index (II) values suggested that ~48% of the identified MGTs can be classified as stable proteins, because of the values less than 40. Also, the grand average of hydropathicity (GRAVY) of rice MGT proteins ranged from -0.054 (in Os01g0601000 from CorA) to 0.789 (in Os06g0715700 from NIPA). Based on our results, three protein types including NIPA, MRS2, and CorA were recognized in MGT proteins based on the protein-specific domain distribution (Table 1).
Phylogenetic relationships and classification of OsMGTs
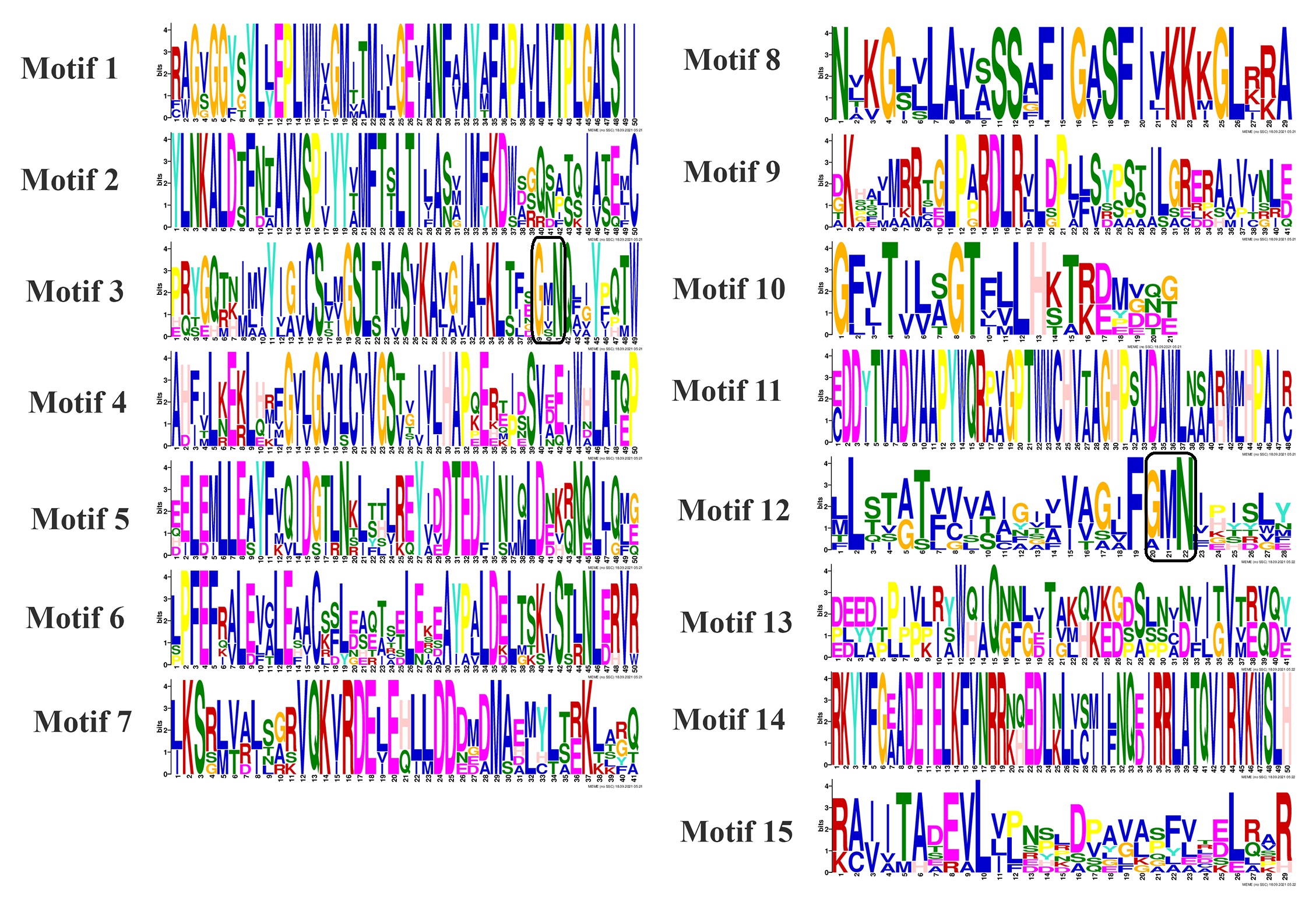
The evolutionary study of the MGT proteins classified them into the MRS, CorA, and NIPA clusters, which demonstrated similar motif patterns in each group (Fig. 1 a and b). As an instance, most of the proteins clustered in the MRS group possessed the motifs 5, 6, 7, 9, and 15 in their structures, while motifs 11 and 13 were detected in all CorA proteins, but a composition of motifs 1, 2, 3, 4, and 10 was predicted in the NIPA group members (Fig. 1 a and b) (Table S1). The conserved transmembrane domains and specific GMN motif at the C-terminal region of MGT proteins were detected in Fig. S1. In addition, the exon-intron structure of OsMGT genes was similar and specific in each cluster. For example, the NIPA group-related members could be suggested as the exon-rich genes with approximately 9-14 exons, while 3-6 coding exons have been predicted in the structure of the MRS-related genes (Fig. 1c). For further evolutionary assay of MGT genes, a phylogenetic tree was constructed using amino acid sequences of MGTs from Oryza sativa, Arabidopsis thaliana, and Zea mays. The analysis result demonstrated that MGT proteins could be clustered in six various groups (Fig. 2). The MRS-related MGTs of these three species were classified into group I, while groups II and III encompassed the CorA proteins; also the NIPA cluster-related members had been grouped into the IV, V, and VI sections. The groups I and VI, both with 32 members, were considered as the largest clades in the phylogenetic tree, while groups IV and III with 3 and 4 members, respectively, had been suggested as the smallest clusters. Moreover, the OsMGTs were present in all groups. According to the phylogeny analysis, the proteins categorized in CorA and NIPA clades were sub-divided into more detailed sub-groups (Fig. 2). According to our findings, it can be suggested that some of the clustered genes may possess recent evolutionary origins, conferring similar functions in the cells.
Genomic distribution, duplication, and synteny analysis of OsMGT genes
All the OsMGT gene family members were successfully mapped onto the 9 out of 12 chromosomes of rice with an unequal distribution (Fig. 3). 7 OsMGT genes were mapped onto the chromosome 1, while only one OsMGT was predicted to be localized on each chromosome 10, 11, and 12. The gene duplication events can generate many genes in various species. Accordingly, 14 segmentally-duplicated/triplicated gene pairs classified in three main groups, including 2, 1, and 11 duplicated gene pairs in MRS, CorA, and NIPA clusters, respectively, were identified in the OsMGT gene family and each group has been demonstrated with colored lines, revealing the paralogous pairs (Fig. 4a). The NIPA group has endured the highest duplicated/triplicated events (Table 2). The paralogous duplicated pairs demonstrated the Ka/Ks ratios from 0.266 to 0.871, estimated to be occurred between 1.9 to 13.8 million years ago (MYA). The Ka/Ks ratios <1 in duplicated gene pairs from the OsMGT family in rice suggested that the genes have been impressed by purifying selection (Table 2). To identify the probable functions of the rice MGT genes, the synteny relationships between the rice, maize, and Arabidopsis genomes had been also investigated. According to the results, all the OsMGT genes showed synteny relationships with their orthologs in the Arabidopsis (~59%) and maize (~46%) genomes (Fig. 4b and c). These wide synteny relations at the gene level can demonstrate the close evolutionary relationships and wide rearrangement events of the rice chromosomes during the genome evolution process.
Gene ontology annotation, post-translational modifications, and promoter cis-elements
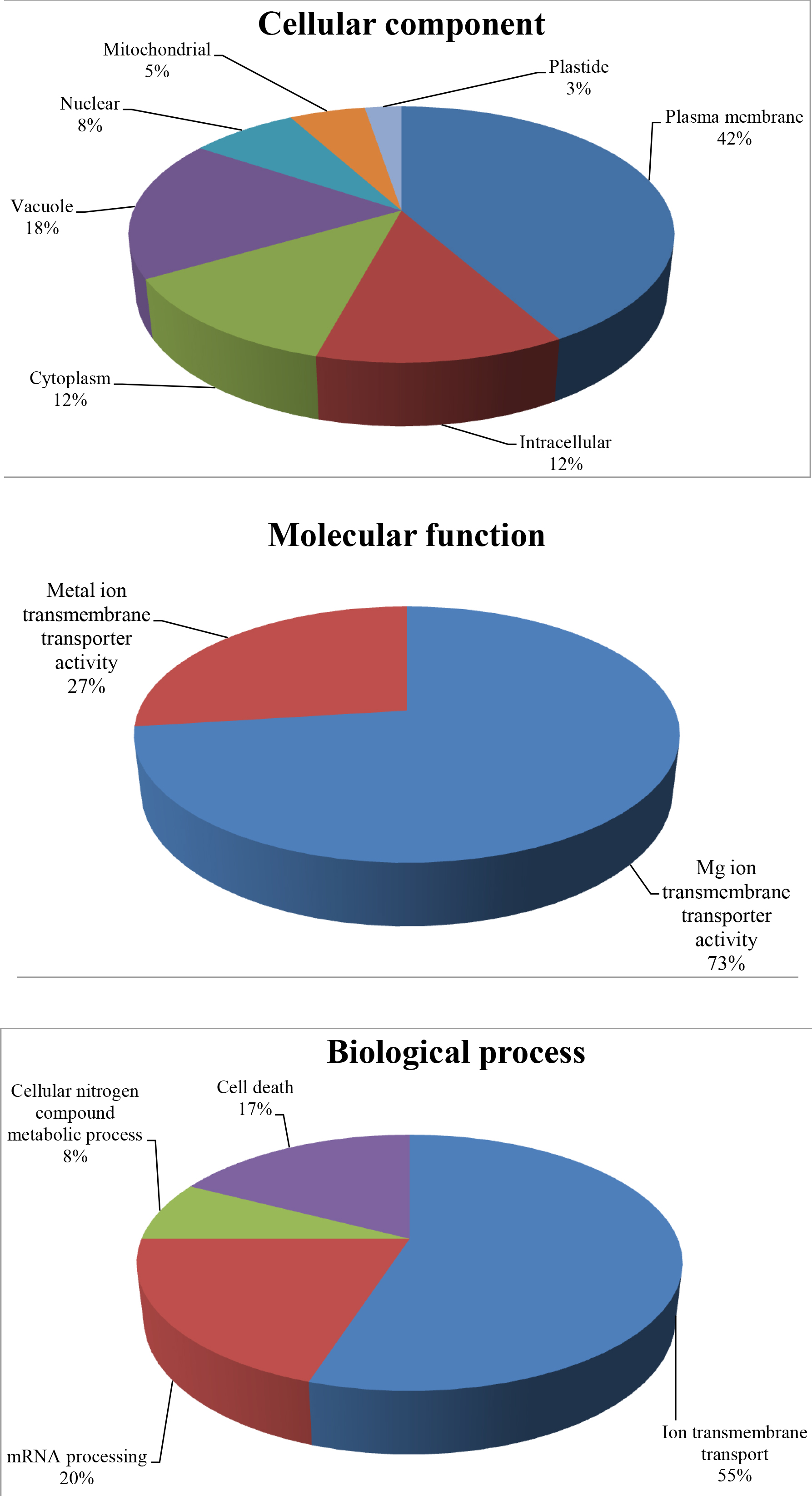
According to the subcellular localization analysis, most of the MGT proteins in rice were localized in the plasma membrane, vacuole, cytoplasm, and intracellular regions, with a fraction of them located into the nuclear, mitochondria, and plastid (Fig. S2). The assessment of the molecular function of OsMGT genes demonstrated that 73% of MGT proteins are engaged in Mg ion transmembrane transporter activity, and 27% of them have a function in transports of other metal ions (Fig. S2). Furthermore, the gene ontology annotation regarding the biological process illustrated that most of the OsMGT proteins are involved in ion transmembrane transport (55%), and the remaining can be engaged in the regulation of mRNA processing (20%), cell death (17%), and cellular nitrogen compound metabolic process (8%) (Fig. S2).
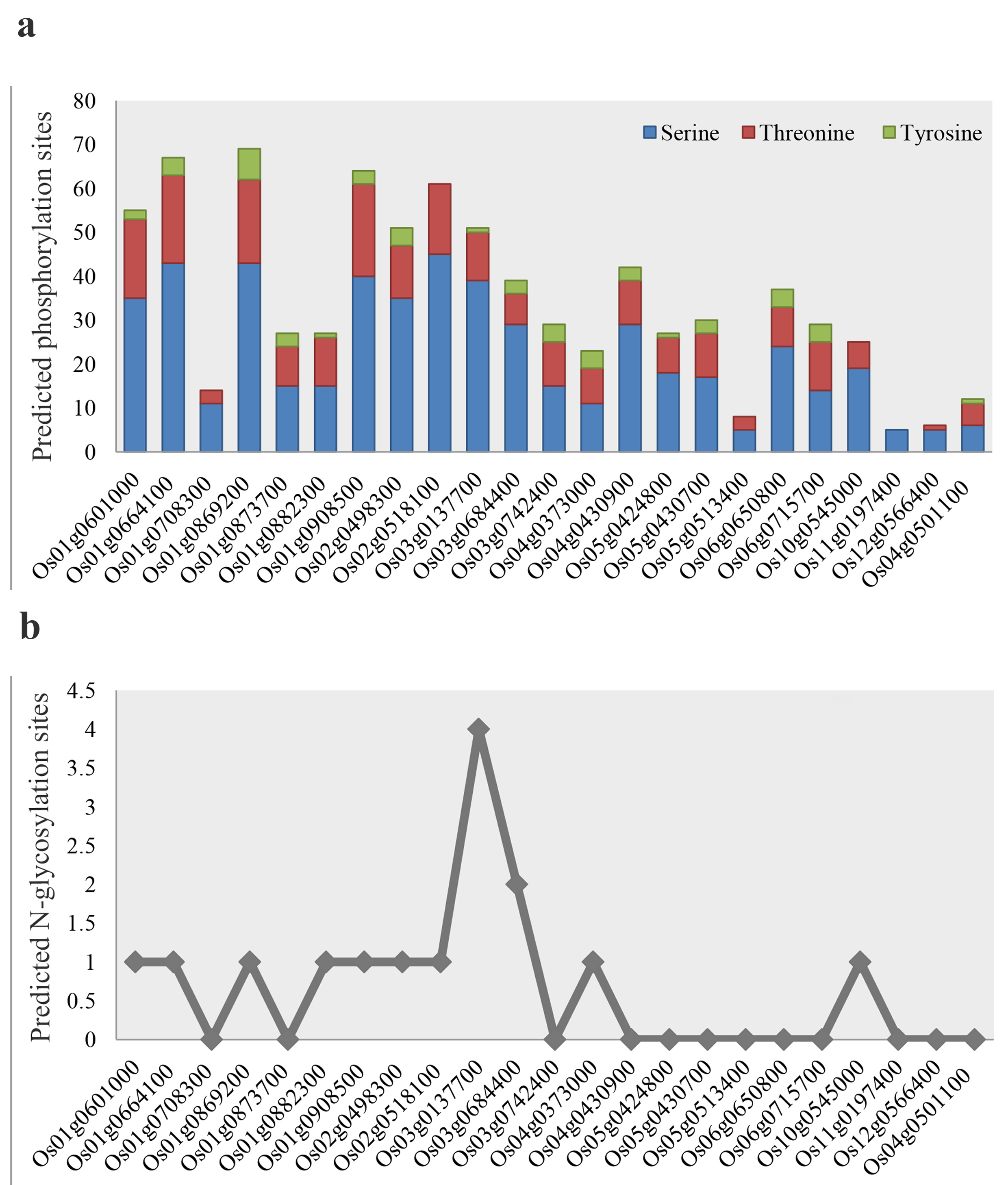
The post-transcriptional phosphorylation sites of OsMGT proteins illustrated a wide variety of phosphorylated serine (S) residues along with some changed threonine (T) and tyrosine (Y) sites (Fig. S3a). About 57% of OsMGTs, such as Os01g0601000 and Os01g0664100 from the CorA clade, Os01g0869200, Os04g0430900, Os01g0908500, and Os03g0137700 from the MRS group, and Os02g0518100, Os02g0498300, and Os05t0424800 belonging to the NIPA category, were predicted as the possible highly phosphorylated MGT proteins in rice. The proteins Os11g0197400 (NIPA), Os12g0566400 (CorA), Os05g0513400 (NIPA), and Os04g0501100 (MRS) were estimated to encompass limited phosphorylated residues in their structures after transcription. Also, most of the phosphorylation events have been occurred in the serine and threonine residues (Fig. S3a). As the important post-transcriptional modification regulating stimuli-induced responses, N-glycosylation sites of Mg transporter proteins were also predicted. Except for about 52% of OsMGTs, for example, Os12g0566400 from the CorA clade, Os01g0708300 in the NIPA group, and Os03g0742400 belonging to the MRS category, the other OsMGT proteins demonstrated some potential N-glycosylated residues at one to four sites (Fig. S3b). Based on the results, Os03g0137700 and Os03g0684400 from the MRS clade were considered as the MGT proteins with many glycosylation sites (2-4 regions), while the other MGTs have been only subjected to one glycosylation during the modification process (Fig. S3b).
In the current assay, the OsMGTs promoter regions in the rice genome were investigated to find the putative cis-regulatory elements. The results demonstrated several kinds of cis-elements for dealing with various phytohormones and abiotic stimulus conditions (Table S2). The promoter common cis-elements, such as the core element TATA-box, and CAAT-box, were distinguished in all OsMGT genes. The ABRE (abscisic acid responsiveness), and TGACG-motif (Methyl jasmonate responsiveness) factors were predicted as the highly occurred hormone-responding cis-elements in approximately all the OsMGT promoters. The light-responsive G-Box and Box 4, anaerobic inducible ARE, WORKY binding site W-box-motif, and stress-responsive MYB elements were detected as the other regulatory cis-elements frequently distinguished in the OsMGTs promoter regions, revealing the significant potential engagement of these ion transporter genes in stimuli coping during the rice life cycle. The MBS (MYB binding site involved in drought-inducibility), LTR (low-temperature responsive), TC-rich repeats (regulating defensive reactions), TGA-element (auxin-responsive), TCA-element (salicylic acid-responsive), and WUN-motif (wounding responsiveness), were identified as the important abiotic/hormone stress-responsive elements significantly predicted in OsMGT genes, especially in the NIPA cluster (Table S2). Regarding our findings, various modulating cis-elements in dealing with phytohormones and environmental stresses were predicted upstream of the majority of OsMGT genes, suggesting the important potential of these ion transporters in rice growth and stress coping.
The secondary structures of the rice Mg transporter proteins
The secondary structures of three types of MGT proteins in rice were predicted using the SOPMA program. The secondary structures of these proteins in rice demonstrated similar patterns in each NIPA, MRS, and CorA cluster, with 23 (in MRS group) to 62% (in NIPA clade) α-helixes, 0 (in NIPA) to 17% (in MRS) β-sheets, 7-24% β-turns (in MRS), and 37 (in NIPA) to 67% (in MRS) coils, (Table 3). The secondary structure α/β patterns in approximately all the OsMGT proteins, especially in the NIPA and MRS clades, were highly similar, illustrating the same functions of these metal ion transporters in the cell. Thus, with significant transmembrane helices (3-9) in the NIPA group proteins as compared with the MRS and CorA clades (0-2), this clade may be effective in rice tolerance to stressful circumstances (Table 3).
The 3D structure modeling, validation, and pocket regions of OsMGT proteins
The MGT protein's three-dimensional structures were predicted under >90% confidence and their potential active sites were also illustrated. The 3D structures of MGT proteins in all MRS, CorA, and NIPA proteins demonstrated a typical frame comprising of various parallel β-turns and α-helixes. According to the results, it can be mentioned that these proteins possess a conserved structure. Despite some determined variations in protein sequences, a significant similarity, especially in the metal ion binding regions and catalytic sites, has been detected in all the MGT proteins. The predicted differences in some regions of these structures can refer to the various roles during transmembrane transport activity under stimuli exposure. Protein structures are exactly associated with the gene functions and also can reflect the phylogenetic relationships. Therefore, although diverse functional groups had been identified in MGTs, it can be proposed that the MGTs from multiple clades share a common catalytic mechanism in ion transmembrane transport and intracellular signaling pathway during stimulus conditions. For assessment of the accuracy and quality of the MGTs 3D models, the Ramachandran plot, as a common analysis for comparison of the rotational angles of proteins, had been employed. According to the program parameters and models validation, the qualities of the MGTs model varied from 87% to 96%, proposing the reliability of the predicted 3D models and the worthy quality (Table 3). For further assessment of the probable errors within the protein models, the ProSA results demonstrated that in each protein model there were the regions with a significant rate of residues with lowest energy, confirming the modeling quality in various parts of these proteins.
The cavities and channel regions in the protein structure can be involved in the regulation of the protein functions in the cells. In this regard, the highest protein channels were estimated to be present in CorA proteins with 8-23 channel regions (Table 3 and Fig. 5). Also the MRS and NIPA group proteins with 4-13 and 3-15 channels, respectively, demonstrated an approximately similar pattern, proposing a similar function in the cells and during multiple stimuli exposure. Based on the results, it can be mentioned that the evolutionary divergence of OsMGTs can adjust the gene's function during several molecular pathways.
Expression analysis of OsMGT genes based on RNA-seq data
The log2-transformed FPKM values obtained from the rice RNA-seq data sets were utilized to comprehend the OsMGT genes functions and transcription patterns of them in various tissues/conditions. All of the OsMGT genes showed the expression level >3 at least in one of the rice tissues (Fig. 6a). Some OsMGTs, such as Os03g0684400, Os03g0742400, Os10g0545000 belonging to the MRS clade, and Os01g0873700, and Os01g0882300 from NIPA, demonstrated the remarkable transcription rates in all the rice tissues, illustrating the critical functions of these Mg transporters during the rice development, which may be adjusted through the regulatory cis-elements in the promoter of the genes (Table S2). Some of the OsMGT genes, such as Os01g0601000 from the CorA group and Os06g0715700 from the NIPA clade, revealed a tissue-specific expression pattern in the embryo and anther tissues, respectively. The Mg transporter genes were strongly expressed in the seed, inflorescence, anther, pistil, callus, and root tissues (Fig. 6b). It can be suggested that these OsMGTs are involved in multiple cellular functions during various developmental stages in rice.
The transcription magnitudes of the rice OsMGT family genes were also investigated in response to multiple abiotic stresses. Six out of 23 OsMGTs, including Os02g0498300, Os06g0715700, Os01g0708300, Os05g0513400 from NIPA, and Os03g0137700, and Os10g0545000 from MRS, were significantly up-regulated in response to all stimuli (Fig. 7a). Also, four Mg transporter genes, including Os01g0869200, Os04g0501100 belonging to the MRS clade, and Os05g0430700, and Os05t0424800 from NIPA, demonstrated the significant down-regulation under stimuli exposure (Fig. 7a). Between the abiotic stimulus conditions, drought stress has induced responses in ~39% of OsMGTs, while under salinity and chilling stresses exposure only ~26 and 9% of OsMGT genes had been induced, respectively (Fig. 8b). Based on our findings, two OsMGTs, namely Os04g0373000 from NIPA and Os12g0566400 from CorA, had been recognized as cold-responsive (Fig. 8b-c). Regarding the significant transcript magnitudes of the NIPA and MRS groups-related genes, they can be considered as the multiple stress dealing Mg transporters during the rice life cycle. These potentials in various OsMGTs may significantly result from the stress-responsive cis-elements present in their promoter regions (Table S2).
Co-expression network of Mg transporters in rice
The OsMGT genes' interactions with other rice genes and their roles in the cells have been investigated through a co-expression network. Based on our predictions, a total of 45 genes had been clustered into the five co-expression nodes in the group A to C in the co-expression network of each main cluster of OsMGTs (Fig. 8). According to the KEGG ontology, 13, 15, and 17 genes were predicted to play roles in the protein ubiquitination (osa04120), amino acids (osa00280) and fatty acid (osa00071) degradations, and phenylpropanoid biosynthesis (osa00940) pathways in the co-expression networks, respectively (Table S3). Between the candidate Mg transporter genes, LOC4327682 (Os01g0708300) as a NIPA-related member in the clade A, demonstrated a significant correlation with an E3 ubiquitin-protein ligase RHB1A (LOC4348088 ), ras-related protein RABA1f (LOC4346744), and ubiquitin-conjugating enzyme E2 4 (LOC4348755), suggesting the important roles in ubiquitin-protein transferase activity and intracellular signal transduction (Table S3). As the other biological pathways modulated by NIPA and its co-expressed genes, protein polyubiquitination, protein phosphorylation, and plant organelle localization can be mentioned that manifest their probable roles in rice growth and stimuli coping. The OsMGT gene LOC4327867 (Os01g0664100), as a CorA candidate, had the functional neighbors, such as F-box protein SKIP28 (LOC4345622), acetyl-CoA acetyltransferase (LOC4326136), ABC transporter B family member 25 (LOC4339984), and disease resistance protein PIK6-NP-like (LOC4344770) in the clade B, which have the functions in developmental and stress-responsive pathways. In the co-expression network related to the candidate CorA, there were also the genes involved in transmembrane transport, amino acid catabolic process, and cellular metal ion homeostasis, which clarified the significant roles during various developmental stages. In clade C, the candidate MRS group-related gene LOC107275769 (Os04g0430900), along with the co-expressed genes MDIS1-interacting receptor-like kinase 1 (LOC4328049), peroxidase 27 (LOC4333262), and lipid-transfer protein 2 (LOC4325636) were predicted to be involved in cellular response to oxidative stress and defense response to abiotic stimuli (Fig. 8 and Table S3). Furthermore, the candidate MRS gene showed an interesting interaction with LOC4329975 and LOC4325035 genes, belonging to the AP2-like ethylene-responsive transcription factors, which have the potentials in the ethylene-activated signaling pathway and stress coping.
{kind=link}
{kind=link}
{kind=link}