The work was organized in two parts, in agreement with the double objective of the study i.e. the detection of plasmid gene markers in LWW and the identification of disinfection / decontamination methods of laboratory cultures achieving the breakdown of antibiotic resistant plasmids.
Detection of the origin of replication of E. coli plasmids in LWW bacteria strains
Wastewater sampling targeted research facilities where the official methods of LLW disinfection are a 20-minute incubation with sodium hypochlorite diluted to 2,4 g.L− 1 sodium hypochlorite (i.e., a 20-fold dilution of a stock solution with 5% active chlorine). Water samples collected from laboratory LWW exhaust ducts and from the main sewage exhaust ducts of research institutes. Bacteria isolates were selected using two criteria. The first selection criterion was the antibiotic resistance profile i.e., the capacity to grow on solid media impregnated with antibiotics frequently used in research laboratories, namely: ampicillin, kanamycin, chloramphenicol, nalidixic acid and spectinomycin. The second criterion was the presence of the origin of replication pBR322 in isolates. This origin of replication is one of the most used in laboratory plasmids. Among 16 antibiotic resistant isolates obtained from 8 LWW samples, 15 bacteria strains presenting the pBR322 origin of replication (Fig. 1) were selected for further analysis (samples L in table S2). Applying the same approach with 36 bacteria strains isolated from research institute sewage exhaust ducts, 10 isolates positive for the pBR322 ori gene were selected for further studies (samples MAS in table S2). Only 1 strain positive for the pBR322 ori gene out of 18 isolates from WWTP samples was selected for further characterization (WWTP-1 in table S2).
The pBR322 origin of replication derives from the EcolE1 plasmid specific of Enterobacteriaceae, especially in Pasteurellaceae (Ares-Arroyo et al. 2018, 2021). Therefore, without adding other origins of replication in these plasmids to broaden their host range, they could normally not replicate in other family bacteria (O’Neil and Bender 1988; (Wu et al. 2010, Ares-Arroyo et al., 2018). However, most isolates positive for the pBR322 ori gene obtained from wastewater samples analyzed in this study were not Enterobacteriaceae. Moreover, the sequence of the pBR322 ori amplicons of most laboratory isolates (L1-6, L9-10, L12 and L15) displayed a 100% homology with that of the commercial synthetic plasmid used as positive control (pBR322 in Fig. 2). The presence of this typical marker of E. coli plasmids in non-Enterobacteriaceae bacteria found in wastewater suggests it could have been acquired through horizontal transfer of laboratory plasmids released undestroyed in the sewage.
This is in line with not only the well described horizontal transfer of plasmid between bacteria, but also its major role in ARGs dissemination (Sorensen et al. 2005 and Pinilla-Redondo et al. 2018). For instance, colE1 plasmids, with the increasingly shift of their gene cargo from bacteriocin to antimicrobial resistance compounds are key-plasmid in ARGs dissemination within Enterobacteriaceae (Ares-Arroyo et al. 2018, 2021 and Dolejska et al. 2018). Another example is the experimental evidence of the transfer of IncHI1A plasmids not only within Enterobacteriaceae but also across other families such as Moraxellaceae, Pseudomonadaceae and Shewanellaceae (Olesen et al. 2022). In line with a possible inter-family transfer of plasmids, in their recent publication Ares-Arroyo and coworkers (Ares-Arroyo et al. 2021) detected the non-conjugative colE1 in non-enterobacteriaceae / -pasteurellaceae bacteria, namely: Pseudomonas, Vibrio, Alivibrio, Photobacterium and Aeromonas.
However, the detection of pBR322 origin of replication in non-Enterobacteriaceae (-Pasteurellaceae) would imply that the carrying plasmid is stable and capable of replication in these bacteria strains. A possible explanation would be that this origin of replication is integrated in a broad host range plasmid possessing multiple replication origins. This seems to be the case in the ColE1 plasmids of certain strains of Aeromonas: beside a putative replication gene they retain the ColE1 origin (pBR322 ori) of replication itself (Ares-Arroyo et al. 2021).
Data about natural transformation of bacteria – including isolates with unexpected pBR322 ori gene in the present study - underlined the necessity to avoid the accidental release of antimicrobial resistant genes in the sewage. This prompted us to lead an investigation of the effectiveness of bacteria neutralization methods routinely used in research facilities to breakdown bacteria DNA (or DNA-bd effect) in treated cells.
Testing the DNA-bd effect of disinfection methods commonly used in the laboratory
As recently pointed out in reviews on this topic (Yoo 2018 and Jones and Joshi 2021) the choice of disinfection methods should rely on in-depth knowledge of their mode of action in view of the increasing number of microorganisms resistant to chemical antimicrobial agents. Accordingly, the present study was focused on the identification of disinfection methods that would be accompanied by a degradation of DNA in treated cells, including plasmid-borne ARG.
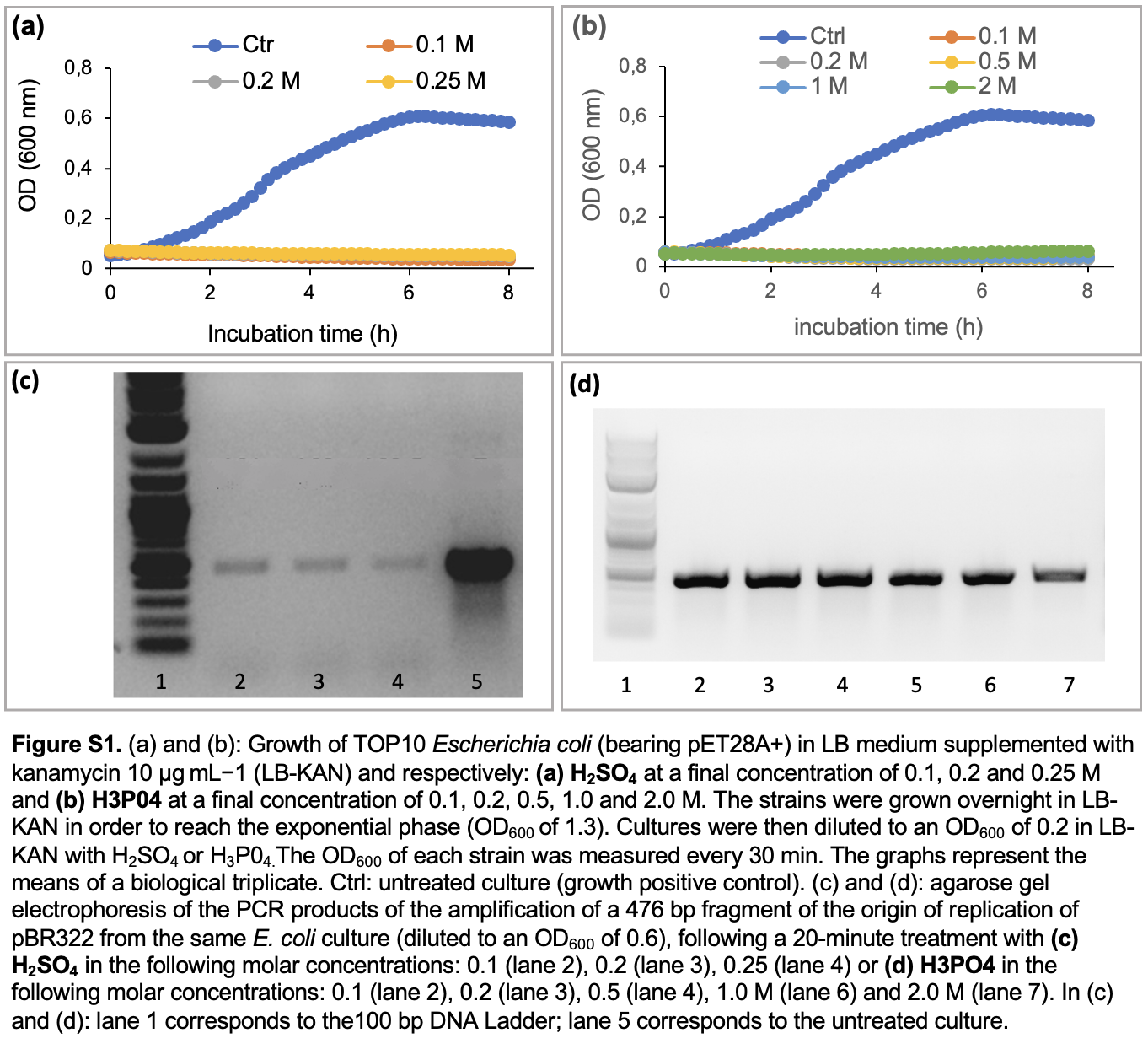
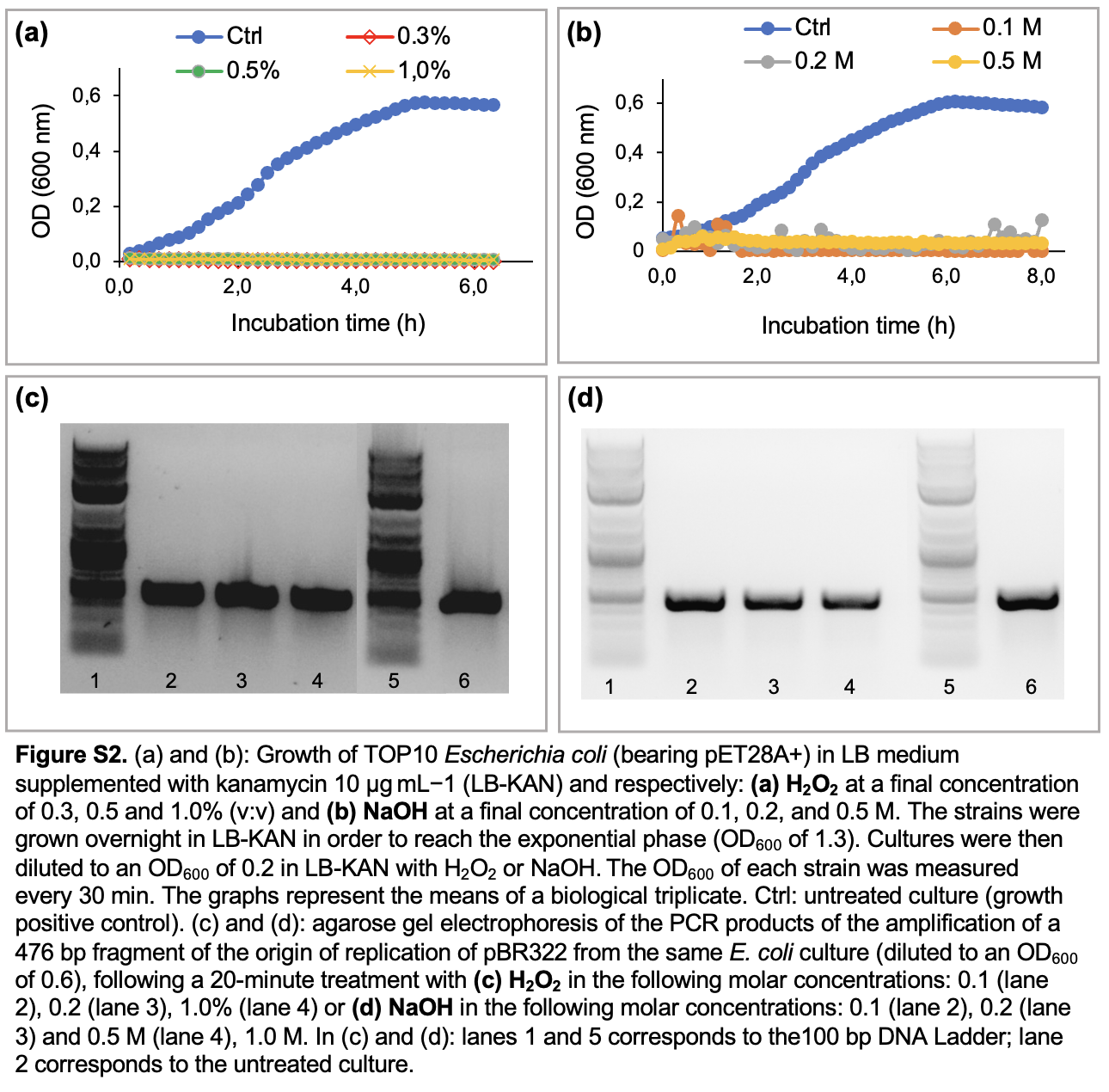
We first investigated the impact of chemical disinfectants on the growth of the model bacterial strain E. coli TOP10 bearing the empty plasmid pET28A+ (biocidal effect). Using the same bacteria culture, the integrity of the plasmid pET28 + following a 20-minute incubation with the same disinfectants was assessed through the PCR detection of one of its gene markers: a 476-bp sequences coding for the pBR322 origin of replication. The tested disinfectants corresponded to the most frequently used biocides in research laboratories, starting with NaClO (stock solution with 5% active chlorine) and a commercial disinfectant referred as P3 (composed of QACS, aldehydes and organic acid), both diluted to a final concentration of 0.5–10% (v:v). As shown in Fig. 3a, bacterial growth was inhibited at all tested concentrations of both biocides (Fig. 3a-b). By contrast, PCR data revealed that only P3 broke down the plasmid DNA. Indeed, in comparison to the untreated culture, the amplification of pBR322 ori gene is reduced in P3 treated cultures (Fig. 3d), while no drastic reduction is seen in PCR products of sodium hypochlorite treated cultures (Fig. 3c). The same experimental protocol was applied with acids, namely: 0.1–0.25 M sulfuric acid (Fig. S1) and phosphoric acid (Fig. S1a-d) which were previously reported as a DNA breakers in a former publication (Kochetkov 1972). Both treatments caused growth inhibition regardless of their respective concentrations (Fig.S1a-b), but sulfuric acid was more powerful to degrade DNA, as seen in PCR results (Fig S1c-d). Sodium hydroxide and hydrogen peroxide also exerted a strong biocidal effect on bacteria (growth inhibition seen in Fig S2a-b), but without perceptible effect on the plasmid DNA integrity (Fig. S2c-d) after a 20-minute treatment. Using quantitative PCR, we quantified the DNA-bd effect of disinfectants with the aim to determine their decimal reduction dose (D-value), reflecting the DNA degradation power. More precisely, D-value corresponds to the quantity of a chemical disinfectant, or the duration of a disinfection method, to decrease the plasmid DNA amount by 90%. For chemical disinfectants tested over a given duration (20 minutes in the present study), the D-value is determined by the linear relationship between the residual quantity of DNA (expressed as log10 value) and the quantity of biocide used (in molar concentration or in %). Using this correlation, the D-values were respectively 0.69 M for sulfuric acid, 4.2 M for phosphoric acid and 3.2% for P3 (Fig. 4).
Regarding physical methods of disinfection often used in laboratories i.e., we compared steam sterilization at 121°C to exposure to ultraviolet light (UVC). Both treatments let to a DNA degradation in a time dependent manner (Fig. S3a-d), confirmed using qualitative PCR (Fig. 4). The D-values (corresponding to the decimal reduction time) were determined as respectively being 25 minutes for steam sterilization at 121°C and 49 minutes for UVC irradiation (Fig. 4c).
Despite an efficient biocide effect of NaClO on bacteria cultures, incubation condition applied in the present study failed to exert a complete DNA-bd effect in dead cells. In agreement with our data, in a previous study (Öncü et al. 2011) the pre-treatment of the transforming plasmids with lower concentrations of active chlorine (between 0.5 and 5 mg.L− 1) had no effect on the efficiency of transformation in vitro suggesting that the treatment had no DNA-bd effect. However, considering the very low concentration used (compared to the conditions applied in the present study i.e. from 25 to 500 mg.L− 1), one could not exclude that NaClO quantities used in their study were below the effective dose. By contrast, a recent study using the same dose than the minimal chlorine concentration used in the present study (i.e. 25 mg.L− 1), it was possible to breakdown bacterial DNA in wastewater, resulting in a 1.7 log10 decrease in the transforming activity of the treated plasmids (Zhang C et al. 2019a). In accordance to this experimental evidence, data obtained by elsewhere indicated that chlorination is an effective method to destruct bacterial ARGs in wastewater treatment plants (Zheng et al. 2017 and (Zhang M et al 2019b). However, the magnitude of the effect of the chlorine-releasing agent (CRA) depended on the antibiotic resistance gene, and the destruction of plasmid DNA was never complete. (Zhang M et al 2019b and (Liu and Hu, 2020). This is not entirely surprising given that, in the process of disinfection, the first targets of CRAs (usually NaClO, ClO2, HCLO and N-chloramines) are the membranes proteins of cells and organelles (Jones and Joshi 2021). Besides oxidizing proteins, CRAs also react with peptides and DNA itself, but proteins of cell membranes and organelles mobilizing a part of free available chlorine (FAC), the resulting DNA-bd effect on plasmid resistant genes is reduced accordingly (matrix negative effect.) As a highly concentrated cell culture was used in the present study (corresponding to usual cultures of E. coli in research laboratories), it cannot be excluded that the poor DNA-bd effect of NaClO results from the interference of cell protein (and other components of the culture medium) on this process, even at high concentration of NaClO (5.000 ppm). In line with this hypothesis sodium hypochlorite is known to be sensitive to the presence of organic material in culture media: a higher concentration is required to achieve disinfection of culture media (2.5000 ppm of available chlorine) and spillages of blood and body fluids (10.000 ppm), compared to surface disinfection (1.000 ppm) ( (Lin et al. 2020)Lin at al. 2020). Another oxidizing agent, peracetic acid (PAA) is mostly used for the surface disinfection of medical material by oxidizing macromolecules (Yoo 2018). Yet, in comparison with chlorine used in the same quantity (i.e. 25 mg.L− 1), PAA is less powerful than chlorine to reduce the transforming activity of treated plasmids (Zhang et al. 2019) and is therefore not recommended for wastewater disinfection.
The commercial disinfectant P3 showed an efficient DNA-bd effect in treated cells. The efficacy of this disinfectant cocktail results from the contribution of its component to destroy macromolecules (proteins and nucleic acids). For instance, QACs – acting as cationic surfactants - destabilize cell membranes (resulting in cell lysis), while aldehydes present in the mix (formaldehyde, glutaraldehyde and glyoxal) also contribute to cell membrane disruption by chemically alkylating the amino (NH2) and sulfhydryl (SH) groups of proteins, as well as the amino groups of nucleic acid bases (e.g. adenine) of DNA and RNA (Lin et al. 2020 and Jones and Joshi 2021). QACs also enhance the bactericidal effect of organic acids present in P3. Therefore, in disinfections procedures developed for laboratories, QACs are attractive biocides not only for the properties mentioned before, but also for their high tolerance towards the contaminating organic matters in cell culture. As a result, they efficiently open the way to aldehydes for DNA breakdown, even in cultures with a high cell concentration (conditions used in the present study).
Regarding acids tested as disinfectants in the present study, phosphoric acid (H3P03), based on its high D-value for DNA destruction. Indeed, a high quantity of phosphoric acid (> 4M) is needed to reduce the DNA amount by only 1 log10. By contrast, the D-value of sulfuric acid determined in the present study (0,7 M) makes it a good disinfectant. However, the routine use of sulfuric acid as a disinfectant being hazardous to user health, as well as to the material integrity, it is also not recommended for routine disinfection.
Among the physical methods of sterilization tested in the present work, sterilization at 121°C appeared a more rapid way to destroy plasmid DNA (with a D-value of 25 min) than UVC (with a D-value of 49 minutes). UVC are routinely used to get rid of microorganism contamination in biosafety cabinets (overnight irradiation), as well as to get rid of DNA contaminating PCR workstations. It is then surprising to see that UVC effects are rather slow to breakdown DNA of bacteria cultures. Yet, recent reports of UV effect on plasmid and ARG integrity are converging to the same conclusion. For instance, with the exception of Zheng and coworkers’ data (Zheng et al. 2017), after UV irradiation of wastewater, there still are antibiotic resistant organisms and ARG in the effluent of a treatment plant (Guo and Kong 2019 and Liu and Hu 2020), due to the matrix effect of liquid waste. The same is true regarding attempts of food sterilizing using UV irradiation (Ballari and Martin 2013).
How to use D-values to design a disinfection method in research laboratories
As mentioned above, the D-value reflects the effectiveness of a disinfection process in terms of biocide concentration or treatment time carried out under a given condition (concentration, sterilization temperature, exposure to UVC, etc.). When comparing disinfection approaches, the smaller the D-value, the more efficient the process. Using D-value, it is possible to assess the Minimum Effective Concentration (MEC) defined as the lowest concentration of the biocide reducing the pathogen titers by 99.99% (Lin et al. 2020). MEC could also be assessed for the DNA-bd effect of chemical disinfectants on the cell DNA.
Under the conditions used in the present study, to reach a 4-log10 reduction of plasmid DNA in treated cells, means using a 20-minute incubation with a final concentration of 13.2% P3 or 2.8 M Sulfuric. The 3-log10 reduction will also be obtained with steam sterilization performed during 100 minutes at 121°C or an UV irradiation performed for 3.3 hours. Practically speaking all those conditions seem not workable, especially for chemicals. Using P3 (and mild acids such as PAA), the D-value could be decreased to a workable one, using higher incubation time (60 minutes instead of 20 minutes, for example). Regarding steam sterilization, the D-value can also be decreased using a higher temperature (134°C). Those options to decrease D-values of DNA-bd effect of disinfection methods will be the next step of our work, including also in vitro biological tests of the transforming capacity of treated samples.
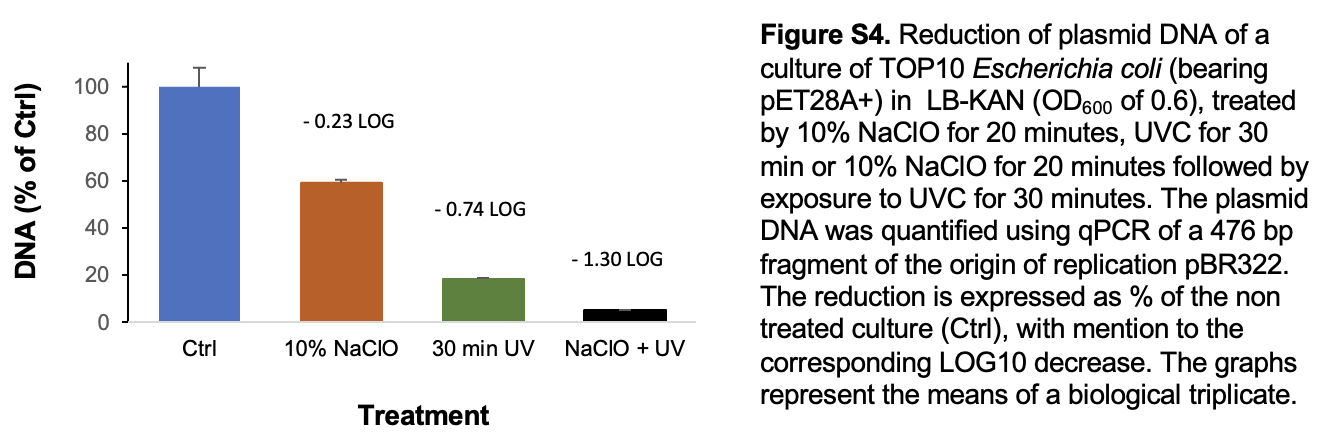
Another alternative is taking advantage of synergies of combined disinfection methods. For instance, preliminary results (Fig. S4) showed that combining 10% sodium hypochlorite with a 30-minute UVC irradiation of a E. coli culture was more effective to breakdown plasmid DNA (1.3-log10 decrease) than the sum of individual effects of NaClO (0,23-log10 decrease) and a 30-min UVC irradiation (0.74-log10 decrease). Though the statistical significance of this preliminary result should be confirmed, it is supported by similar synergies reported for the combination UVC and chlorine to reduce the conjugative transfer (Lin et al. 2016) or to degrade of ARG (Liu and Hu 2020). The synergy could be explained by a better access of UV to the DNA of the bacteria destroyed by chlorine, combined with a reduced capacity of the DNA repair systems of the bacteria under double treatment.
Considering the increasing evidences about the capacity of microorganisms to develop resistance to biocides (Russel 2002, 2003), which in turn could lead to increased resistance to antibiotics (Maillard 2018 and (Nicolae Dopcea et al. 2020), methods combining a chemical and physical disinfections seem to be the appropriate way to limit the resistance development, as it will avoid using increased amounts of chemicals to breakdown plasmid DNA. The well documented resistance to QACs (Zou et al. 2004), associated to several resistance genes (sugE(c), ydgE/ydgF, mdfA, emrE and qacEDelta1) and the demonstration of the horizontal transfer of one of them (qacEΔ1) (Jiang at al. 2017 and Han et al. 2019) is especially alarming. As a result, two recommendations were recently issued regarding the use of chemical disinfectants. Firstly, biocides should only be used in their minimal inhibitory concentration (MIC) (Vijayakumar and Sandle, 2019). However, as seen in the present study, in MIC doses disinfectants would not completely destroy plasmid DNA. The second recommendation is an urgent call to apply the current policy about antibiotic use to antiseptics / biocides, in order to preserve their biocidal benefits (Nicolae Dopcea et al. 2020). Both recommendations point out the need to develop education programs on the appropriate use of disinfectants in research laboratories, especially in low containments (Biosafety level 1 and 2 laboratories) where the low risk of acquiring an infection on the workplace could lead to an underappreciation of the high risk of dissemination of ARG – and therefore to contribute to the global antibiotic resistance crisis – due to accidental release of undestroyed plasmids in sewage.
{kind=link}
{kind=link}
{kind=link}
{kind=link}