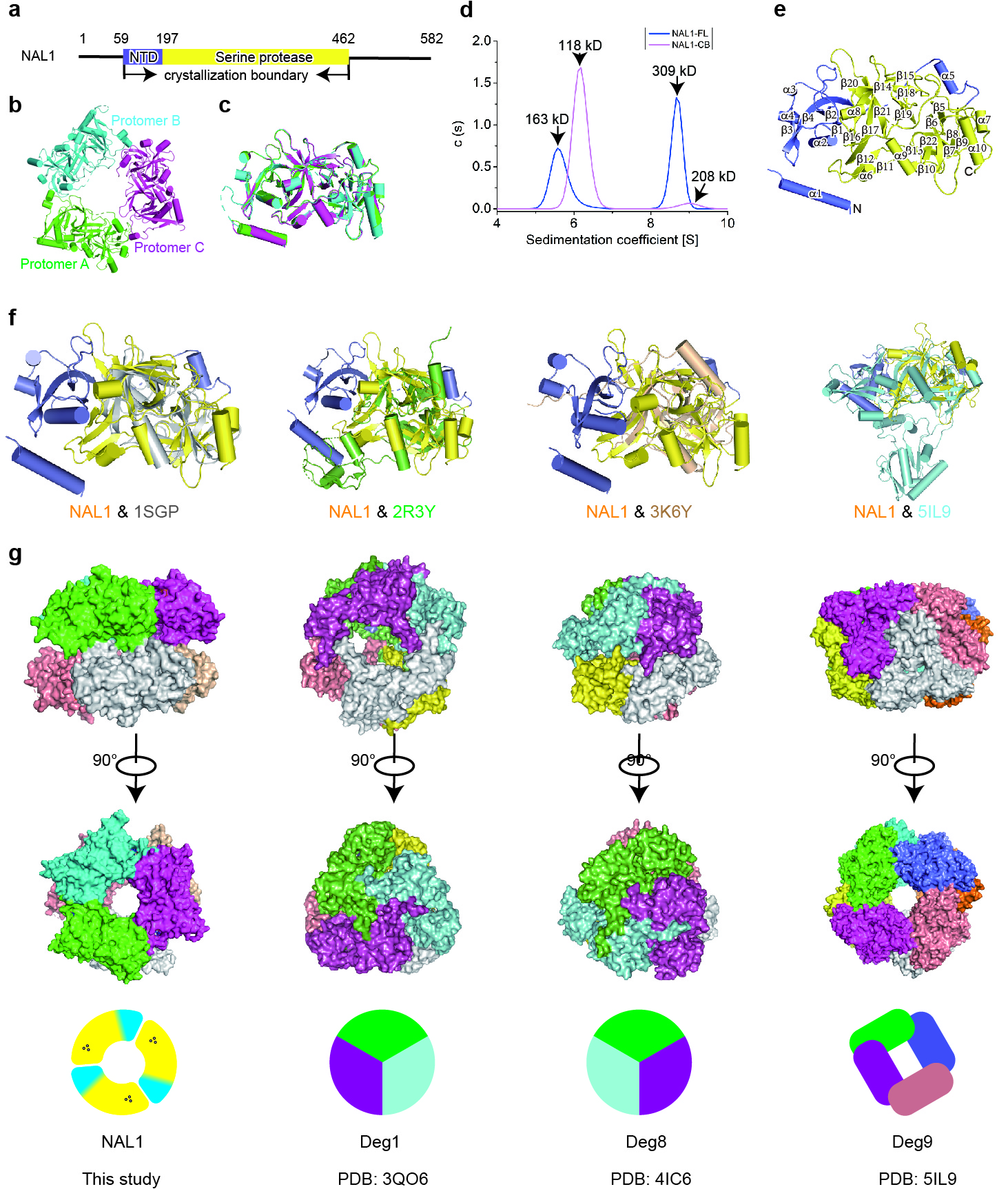
Crystal structure determination of NAL1 hexamer
In the past decade, several studies have demonstrated the pleiotropic roles of NAL1 in agronomic traits, and it has been identified as a valuable gene in rice breeding9,12,15. In order to unveil NAL1 pleiotropic mechanism, we performed protein structure analysis since sequence alignment of NAL1 against known proteins could not predict a reliable biochemical function. The crystal structure of NAL1 (residues 59-463) was determined at a resolution of 2.6 Å (Fig. 1a, Extended Data Fig. 1a and Extended Data Table 1). Electron density of residues 78-84, 175-183, and 407-412 was undetectable, which might be due to their flexibility in solution. NAL1 was trimeric in one unit cell (Extended Data Fig. 1b), and NAL1 hexamer was assembled from two symmetric neighboring unit cells (Fig. 1a). The protomers of NAL1 shared almost identical architectures without apparent structural variation (Extended Data Fig. 1c). To further validate the oligomeric state of NAL1, analytical ultracentrifugation (AUC) experiments were performed. The results showed that the full-length NAL1 was mainly in a hexameric state, which was consistent with the crystallographic analysis (Extended Data Fig. 1d).
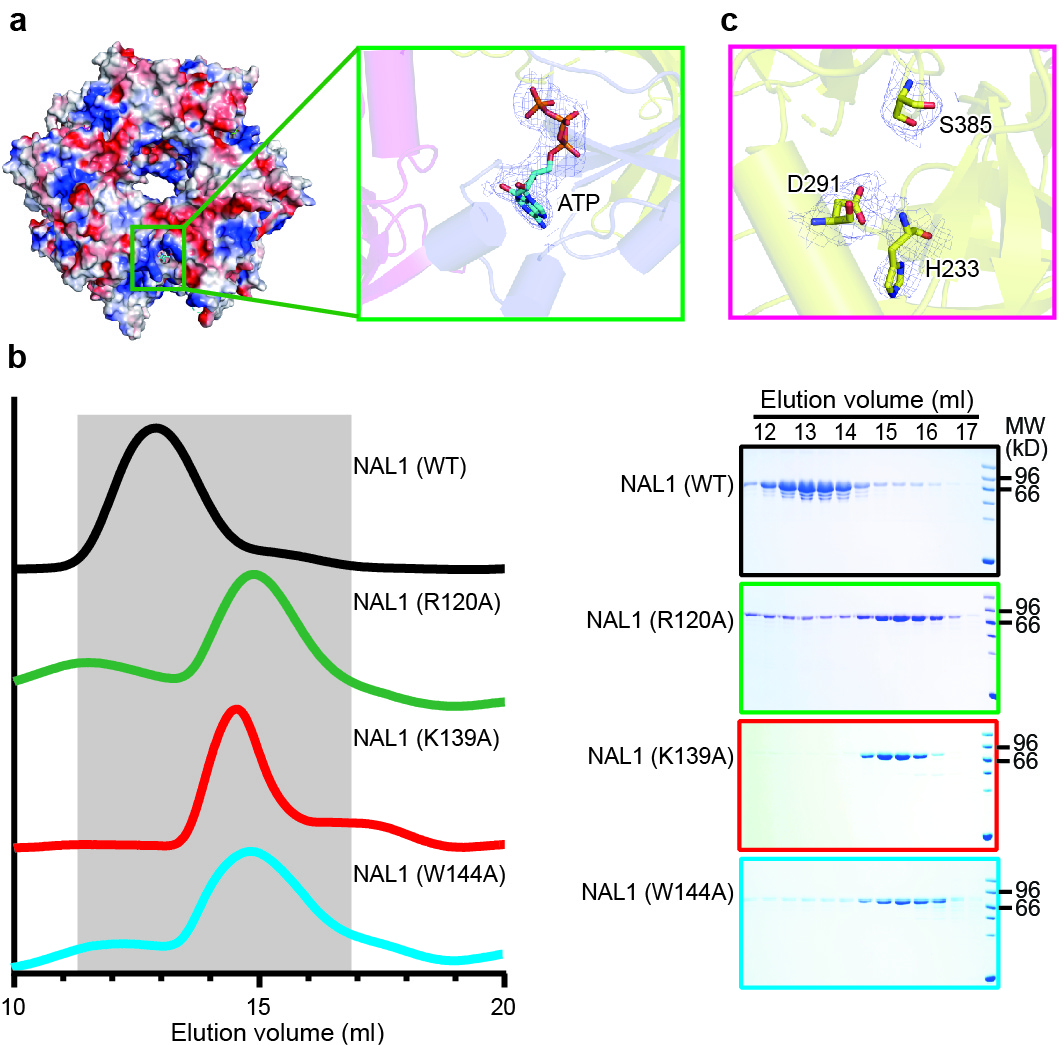
Dali (a structure comparison server) search revealed that NAL1 was a serine protease according to the structural similarity (Extended Data Fig. 1f, and Extended Data Table 2). Thus, each NAL1 protomer was divided into two segments: N-terminal domain (NTD, residues 59-197) and C-terminal serine protease domain, consisting of 10 α-helices and 22 β-sheets (Extended Data Fig. 1a, e). Within the NTD, an ATP molecule was found in a pocket surrounded by a3, a4, b4, and a loop (Fig. 1b, Extended Data Fig. 2a), and this pocket was stabilized by a positively charged network consisting of residues R120, R122, K123, K139, and K142 (Fig. 1c). This ATP molecule expelled a3 from one protomer to another, thus leading to a cation-p interaction between W144 and H248 from the neighboring protomers, eventually mediating the formation of a trimer. Our results showed that the oligomerization was abolished by W144A point mutation, further validating role of the cation-p interaction in oligomerization (Extended Data Fig. 2b). Further, we investigated the effect of the residues surrounding ATP on NAL1 oligomerization. The mutation of either R120 or R139 into alanine disrupted the oligomerization (Extended Data Fig. 2b), suggesting that ATP could mediate the oligomerization of NAL1.
The C-terminal serine protease domain of NAL1 consisted of two perpendicular six-stranded β-barrel subdomains, and the putative catalytic triad was composed of H233, D291, and S385, which were located in the crevice between the two lobes (Fig. 1d, Extended Data Fig. 2c). We noticed that the natural single nucleotide variation (A-to-G substitution) of NAL1 occurred exactly in the catalytic triad, resulting in the conversion of His233 to Arg233 (H233R), which was found to be associated with the pleiotropic traits11,17. In this study, we designated the two alleles of NAL1 as NAL1A (H233) and NAL1G (R233), respectively. To genetically examine the functional differences between the alleles, we constructed two complementary lines NAL1A-COM and NAL1G-COM in the NAL1-knockout mutant (nal1-cri) background generated by CRISPR-Cas9 technique. Both complementary lines partially recovered phenotypes of nal1-cri, but the NAL1A-COM line showed significantly stronger phenotypes than the NAL1G-COM line, including wider flag leaf, higher plant height, larger root, larger panicle, and higher grain yield (Fig. 1e-j). These results suggested that the NAL1A was a genetically stronger allele, and that the H233 in the catalytic triad might be a critical residue for NAL1 functions.
NAL1 physically interacts with OsTPR2
To unveil the underlying mechanism of NAL1’s pleiotropic functions, immunoprecipitation in combination with mass spectrometry (IP-MS) was used to identify NAL1-interacting proteins. Of the detected 31 proteins (Extended Data Table 3), 3 TPR family proteins (OsTPR1/2/3) were selected for further research since they regulate multiple growth and development processes28,32.
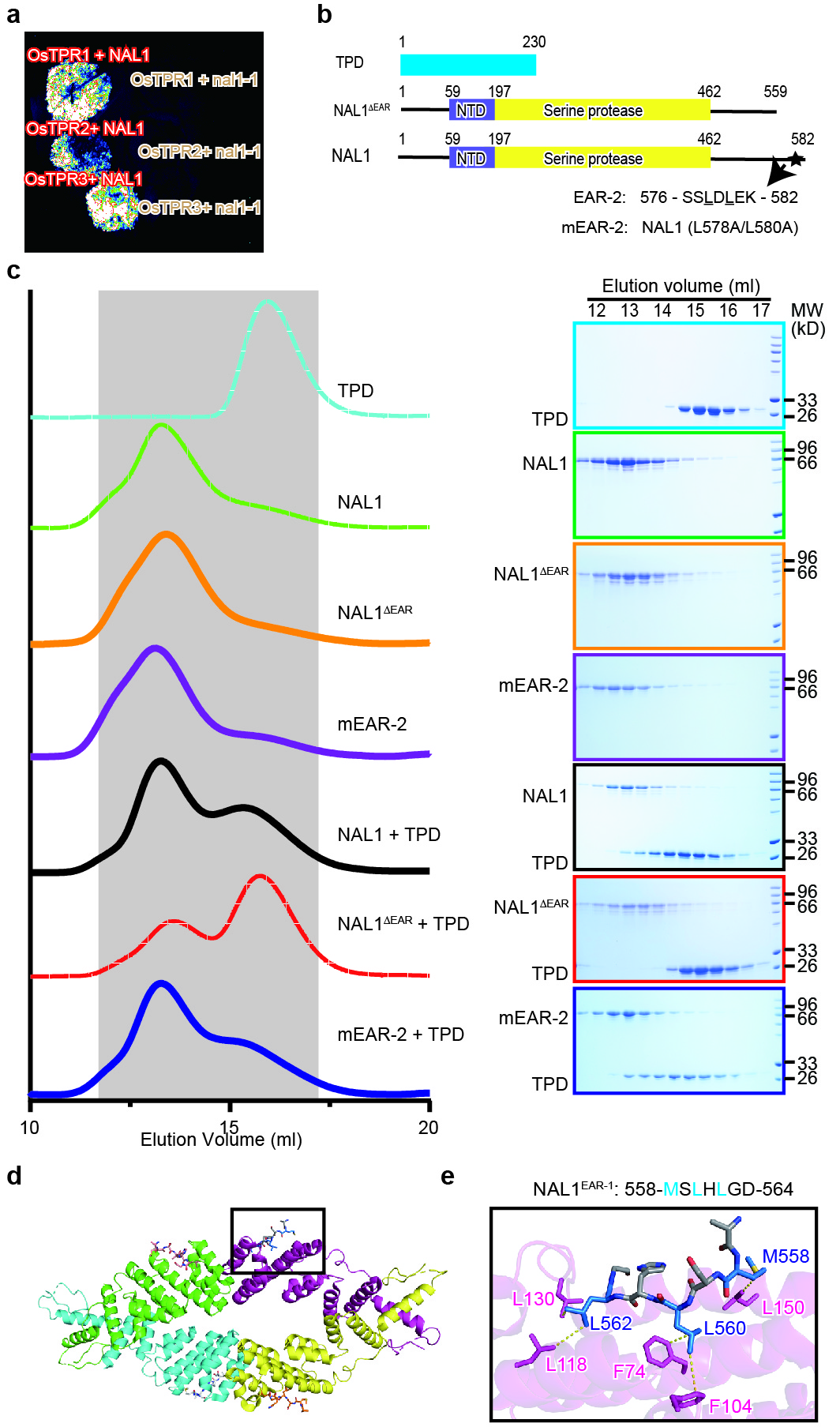
To verify the interaction between NAL1 and OsTPR family proteins, we performed luciferase complementation imaging (LCI) assays in Nicotiana benthamiana leaves. The previously reported NAL1 mutant (nal1-1) with 10-AA deletion in the serine protease domain19 was used as a negative control. We found that NAL1, rather than nal1-1, interacted with all the three rice TPR proteins (Extended Data Fig. 3a). OsTPR2 was selected for further investigation since this gene has been reported to be involved in regulating multiple developmental and agronomic traits33-36. Bimolecular fluorescence complementation (BiFC) assays showed that YFP fluorescence signal was detected in the cells co-expressing OsTPR2-nYFP and NAL1-cYFP, but not in the cells co-expressing OsTPR2-nYFP and nal1-1-cYFP (Fig. 2a). In addition, NAL1-GFP was co-immunoprecipitated with OsTPR2-Flag when they co-expressed in tobacco leaves (Fig. 2b). All above results suggested the interaction between NAL1 and OsTPR2.
OsTPR2 was divided into N-terminal TPD domain and C-terminal WD40 domain according to previous reports32,37. LCI assay was performed to further determine which domain of OsTPR2 interacted with NAL1. The results showed that TPD, rather than the WD40 domain, interacted with NAL1 (Fig. 2c), implying that N-terminal TPD domain mediated the interaction between NAL1 and OsTPR2. Previous reports show that TPD interacts with proteins via an EAR-motif (LXLXL)38-41. We detected two EAR-like motifs (EAR-1: 558-MSLHLGD-564; EAR-2: 576-SSLDLEK-582) in the C-terminus of NAL1 (Fig. 2d, Extended Data Fig. 3b). To further determine whether these two EAR-like motifs mediated the interaction between NAL1 and OsTPR2, a truncated NAL1 with the two EAR-like motifs deleted (residues 1-559) was constructed. Gel filtration assays showed that the deletion of two EAR-like motifs completely abolished the interactions (Extended Data Fig. 3c). Point mutations at the EAR-1 motif disrupted their interaction (Fig. 2d), whereas those at the EAR-2 motif did not (Extended Data Fig. 3c), suggesting the key role of EAR-1 in the interaction between NAL1 and OsTPR2. To further decipher the interaction mechanism between NAL1EAR-1 and OsTPR2, we modeled a OsTPR2-NAL1EAR-1 complex structure, in which L560 and L562 of NAL1EAR-1 exhibited a hydrophobic interaction with F74, F104, L118, L130 of OsTPR2 (Extended Data Fig. 3d, e). Taken together, NAL1 C-terminal EAR-like motif and OsTPR2 N-terminal TPD domain jointly mediated the interaction between NAL1 and OsTPR2.
NAL1 promotes degradation of OsTPR2
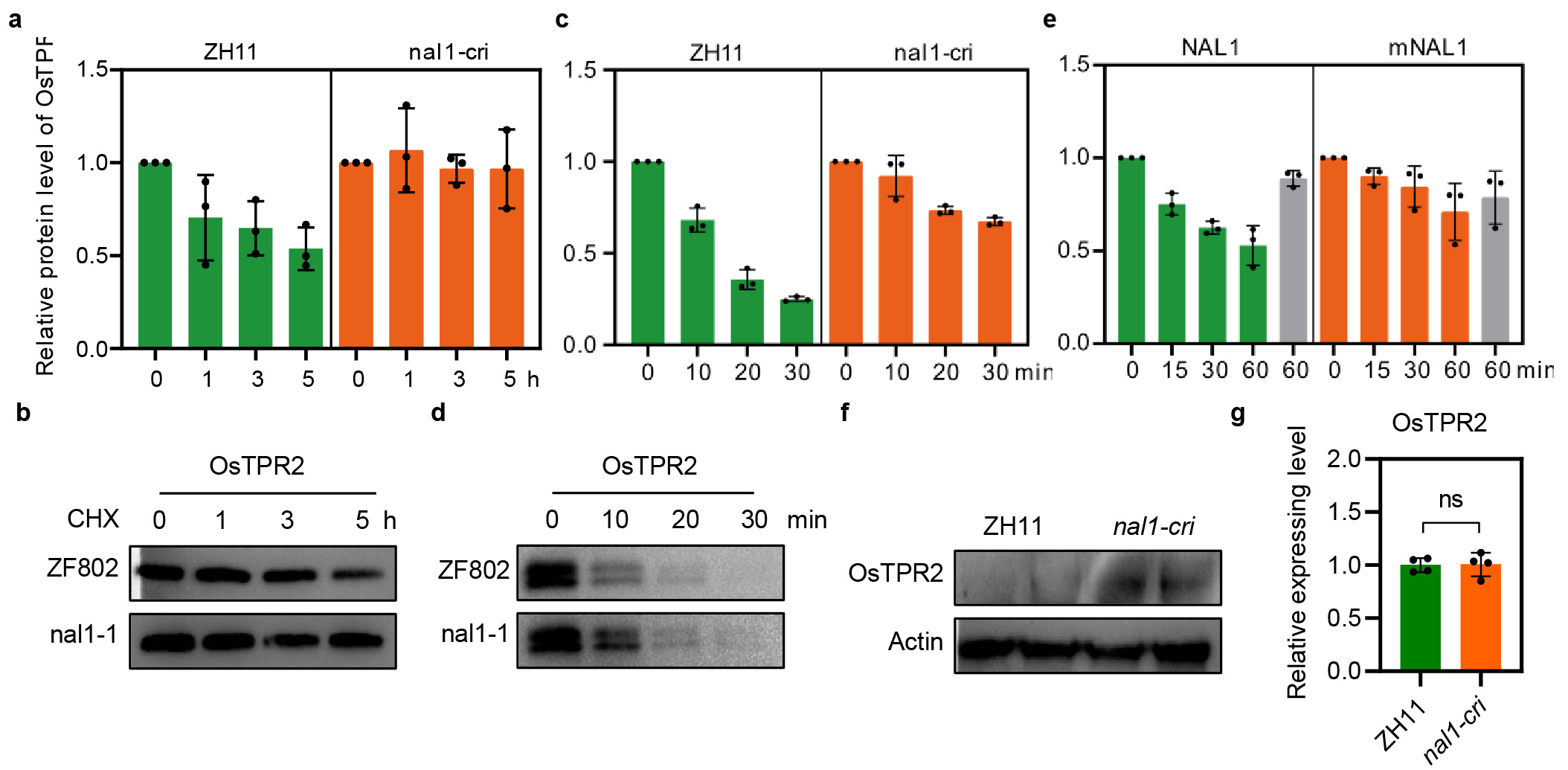
To test whether NAL1, a structurally distinct serine protease, is capable of degrading OsTPR2, in vivo protein degradation assays were performed using rice protoplasts. The results showed that OsTPR2 was hardly degraded in nal1-cri protoplasts, whereas ~50% OsTPR2 was degraded within 5 h in the wild type (ZH11). As a negative control, GFP protein exhibited no change in nal1-cri and ZH11 protoplasts (Fig. 2e, Extended Data Fig. 4a). Moreover, the same assays were performed in the nal1-1 mutant and its corresponding wild type (rice cultivar ZF802)19, and similar results were observed (Extended Data Fig. 4b). Overall, our results showed that NAL1 could facilitate the degradation of OsTPR2 in vivo.
Subsequently, recombinant 6×His-tagged OsTPR2 was used as substrate to investigate its degradation by NAL1 in cell-free extracts from nal1-cri and wild type ZH11 plants. Compared to ZH11, the degradation of OsTPR2 was delayed in the nal1-cri extract (Fig.2f, Extended Data Fig. 4c). Similar results were observed in the assays of cell-free extracts of nal1-1 mutant and wild type ZF802 plants (Extended Data Fig. 4d). These results further suggested that NAL1 could promote OsTPR2 degradation. To further confirm whether NAL1 functions biochemically as a serine protease, different protease inhibitors were added into ZH11 cell-free extracts to examine the degradation of OsTPR2. The results showed that all the 3 serine protease inhibitors phenylmethanesulfonyl fluoride (PMSF), aprotinin, and leupeptin could almost abolish OsTPR2 degradation. In contrast, the metalloprotease inhibitor bestatin, the aspartic protease inhibitor pepstain, or the cysteine protease inhibitor E-64 could not inhibit OsTPR2 degradation (Fig. 2g, h). These results strongly suggested that NAL1 was a bona fide serine protease.
To examine the importance of the catalytic triad to the proteolytic activity of NAL1, wild type NAL1 and the triad-mutated NAL1 (mNAL1) were incubated with OsTPR2 in vitro, respectively. The results showed that wild type NAL1 degraded ~50% OsTPR2 within 1 h, whereas mNAL1 exhibited highly compromised degradation activity (Fig. 2i, Extended Data Fig. 4e). To further reveal the function of the catalytic triad of NAL1 in plants, we examined the proteolytic activity in the previously reported near isogenic lines NIL-IR and NIL-ZS containing the alleles NAL1A and NAL1G, respectively17. In vivo and cell-free protein degradation assays both showed that NAL1 protein from NAL1A allele had stronger proteolytic activity than that from NAL1G (Fig. 2j, k). In addition, the OsTPR2 abundance quantified by the anti-OsTPR2 antibody was significantly higher in nal1-cri than in ZH11 (Extended Data Fig. 4f), while the OsTPR2 mRNA level showed no difference between nal1-cri and ZH11 (Extended Data Fig. 4g). These results suggested the importance of the catalytic triad to the proteolytic activity of NAL1.
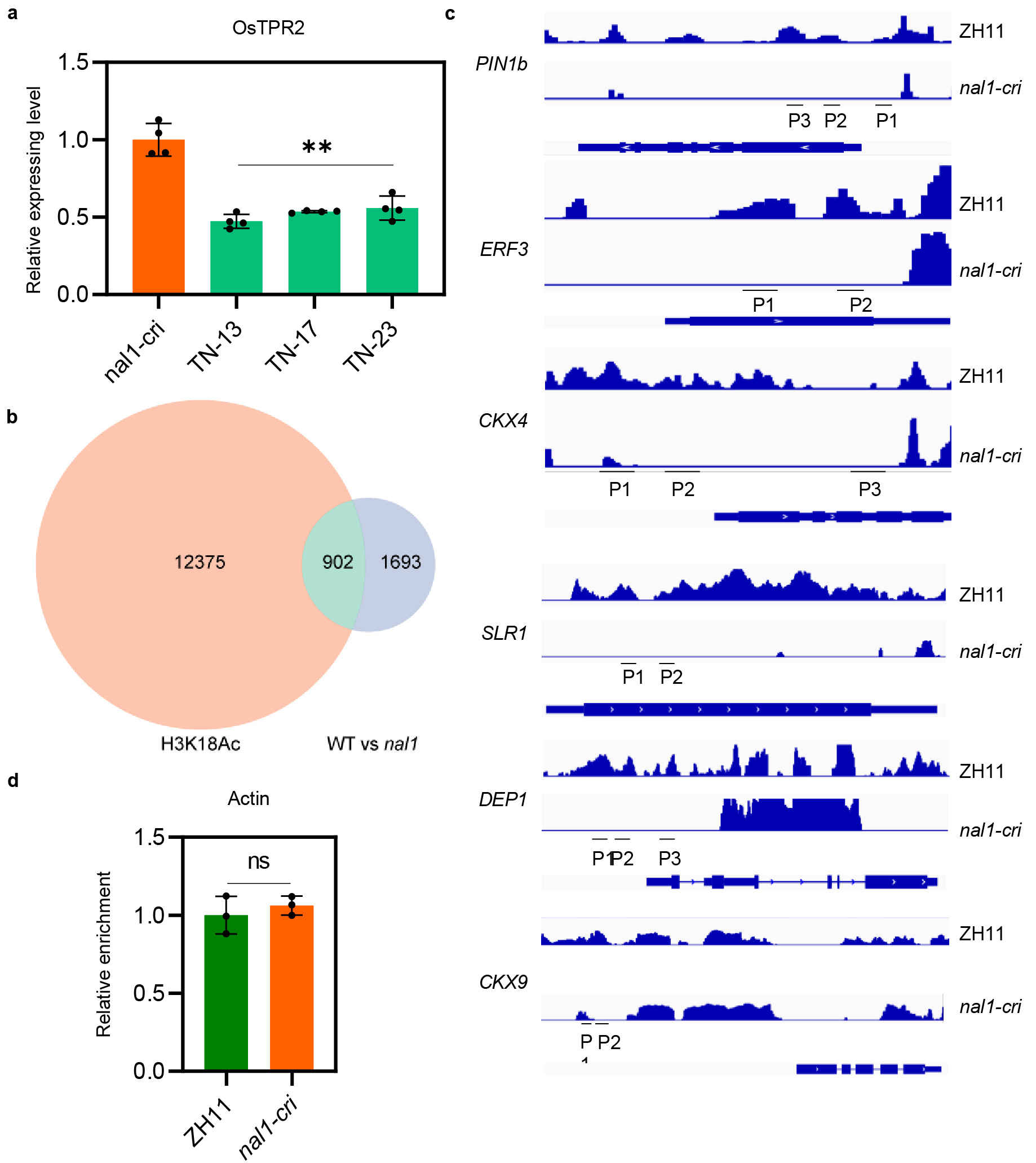
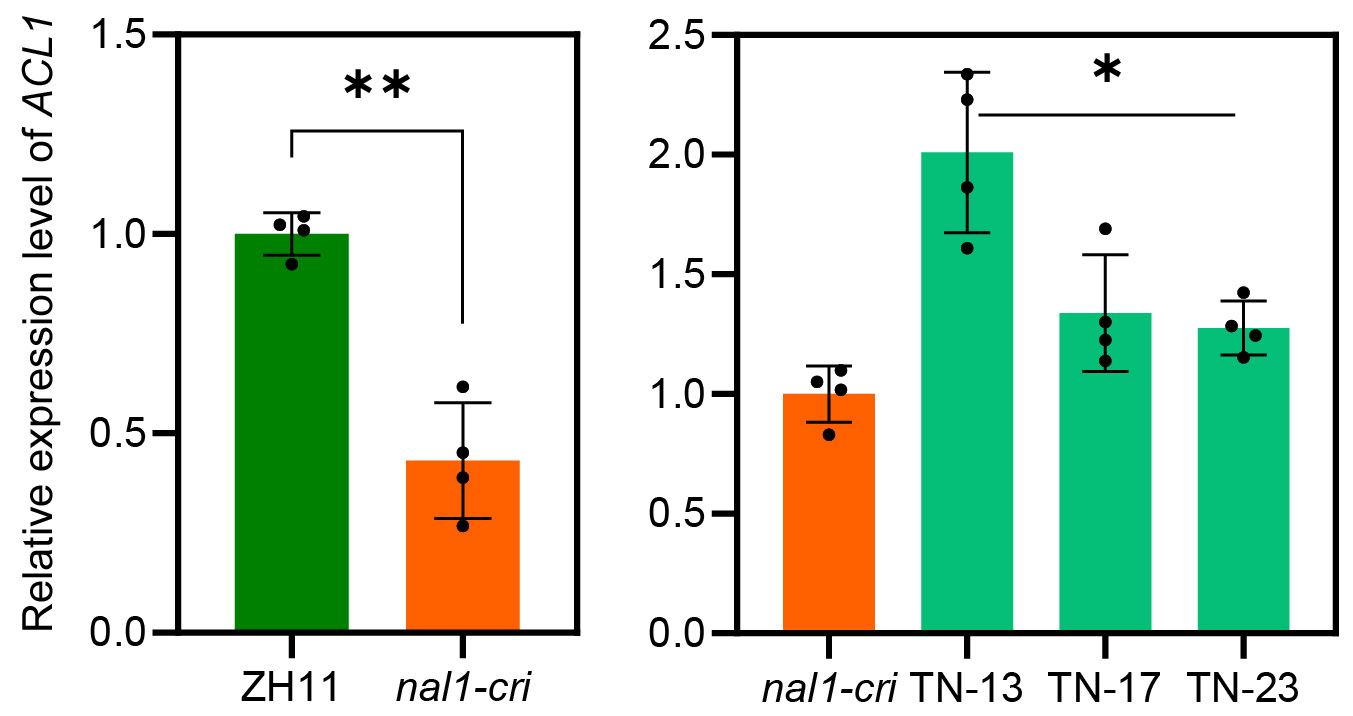
To clarify the genetic interaction between NAL1 and OsTPR2, we constructed OsTPR2 RNA interference (RNAi) lines in the nal1-cri background. The expression level of OsTPR2 in three positive OsTPR2-RNAi/nal1-cri lines was decreased to approximately half of that in nal1-cri (Extended Data Fig. 5a). Compared to those in nal1-cri, plant height, flag leaf width, root dry weight, and grain yield were significantly increased in the three OsTPR2-RNAi/nal1-cri lines (Fig. 3a-f). These results suggested that the pleiotropic function of NAL1 genetically depended on OsTPR2, which was consistent with the biochemical relationship between NAL1 and OsTPR2.
NAL1 exerts pleiotropic function by alleviating OsTPR2-mediated hormone signaling pathway suppression
TPL/TPR family proteins have been reported to regulate hormone signaling pathways by forming complexes with repressors and transcription factors, thus affecting histone acetylation levels of downstream target genes in Arabidopsis26-28,32,34,38,39. To determine whether the rice homologs share the same mechanism, we investigated the potential complex formation from OsIAA3/17 (identified by a yeast-two-hybrid screening of OsTPR2), OsARF25 (OsIAA3/17-interacting transcription factor in auxin signaling pathway)42,43, and OsTPR2 by in vitro pull-down assays. Meanwhile, the repressor D5334 and its corresponding transcription factor IPA144 in SL signaling pathway were selected for examining their interactions with OsTPR2. The results showed that GST-tagged OsARF25 (or IPA1) was co-eluted with 6×His-tagged OsIAA3/17 (or D53) and StrepII-tagged OsTPR2, but not with StrepII-tagged OsTPR2 alone (Fig. 4a-c), suggesting that OsTPR2 might participate in the regulation of hormone signaling by forming a TF-adaptor-TPR complex like their homologs in Arabidopsis. Since NAL1 promoted OsTPR2 degradation, we speculated that NAL1 might affect histone acetylation, thus alleviating the OsTPR2-mediated suppression of genes in the hormone signaling pathways. To test the speculation, the histone acetylation levels in ZH11 and nal1-cri were examined. H3Ac, H3K9Ac, and H3K18Ac levels were lower in nal1-cri than in ZH11, with H3K18Ac exhibiting the most significant difference (Fig. 4d). Furthermore, a total of 13277 genes were found to have a lower H3K18Ac level in nal1-cri than in ZH11 by ChIP-seq analysis. Based on the published RNA-seq data of nal1 mutant and its wild type20, 902 genes whose expressions were down-regulated in the nal1 mutant20 also showed decreased H3K18Ac levels (Extended Data Fig. 5b, and Extended Data Table 4), further supporting that NAL1 fulfilled its pleiotropic functions by alleviating the OsTPR2-mediated gene expression suppression.
To further clarify how the NAL1-OsTPR2 module regulated gene expression, we investigated OsARF25-targeted genes (PIN1b, ERF3 and CKX4) in auxin signaling pathway and IPA1-targeted genes (SLR1 and DEP1) and CKX9 (affected by D53, but not the target gene of IPA1) in SL signaling pathway. The genome browser data showed that H3K18Ac level of these genes was significantly lower in nal1-cri than in ZH11, which was confirmed by H3K18Ac-ChIP qPCR assays (Fig. 4e, f, Extended Data Fig. 4c, d). We examined the effects of NAL1 on the expression of these genes. As expected, these genes showed lower expression in nal1-cri than in ZH11 (Fig. 4g). In the double mutant OsTPR2-RNAi/nal1-cri lines, down-regulating OsTPR2 partially alleviated the expression suppression of above-mentioned genes, compared to nal1-cri (Fig. 3g). These results jointly implied that NAL1 positively modulated histone acetylation and gene expression by degrading the co-repressor OsTPR2.
NAL1 and its homolog genes in other crops are highly breeding-valuable
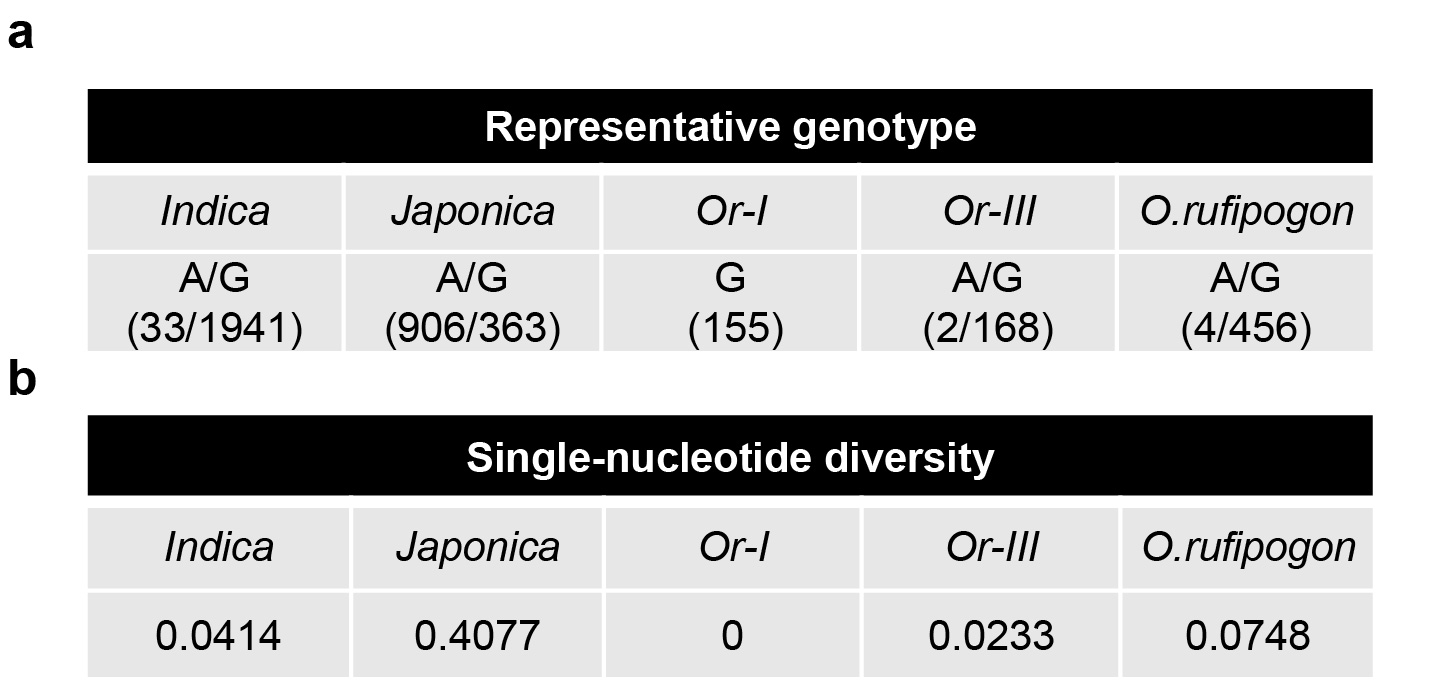
In previous studies, the natural variation of NAL1, mainly two haplotypes corresponding to the alleles NAL1A and NAL1G, is genetically detected in cultivated Asia rice varieties10,11,15. The difference of their protease activity (Fig. 1, 2) was in accordance with their biological function in vivo. It has been reported that the stronger allele NAL1A is derived from the weaker allele NAL1G in the ancestral population and has undergone artificial selection8,18,45, but its detailed origin route is still unknown. Therefore, we analyzed whole-genome sequencing data of a large panel of accessions including 3243 O. sativa varieties and 460 O. rufipogon accessions46,47. NAL1A exists in 71.39% of japonica rice varieties, but only in 1.67% of indica rice varieties, indicating that the NAL1A might be population-specific. Indica and japonica have been reported to be descended from common wild rice Or-I and Or-III, respectively46. We noticed that NAL1A existed only in 2 Or-III accessions, but NAL1G existed in all the Or-I accessions (Fig. 5a, Extended Data Fig. 6a), implying that NAL1A in japonica rice was originated from Or-III wild rice. Single-nucleotide diversity analysis showed that the diversity of japanica was almost 20 folds higher that of Or-III, but indica did not significantly differ from Or-I (Extended Data Fig. 6b), indicating that NAL1A might undergo artificial selection during japanica rice domestication, which is consistent with previous report45.
Since almost all indica rice varieties contain the weak allele NAL1G, we wondered whether the functionally strong allele NAL1A has potential in indica rice breeding. To this end, we introduced the NAL1A allele into Huang-hua-zhan (HHZ), an elite indica variety widely cultivated in China, to replace its NAL1G allele. Morphological traits including plant height, flag leaf width, root and panicle size were significantly improved in the near-isogenic line HHZ-NAL1A, compared to those in HHZ. As a result, grain yield per plant was significantly increased (Fig. 5b-g), indicating promising application value of the NAL1A allele in indica rice breeding.
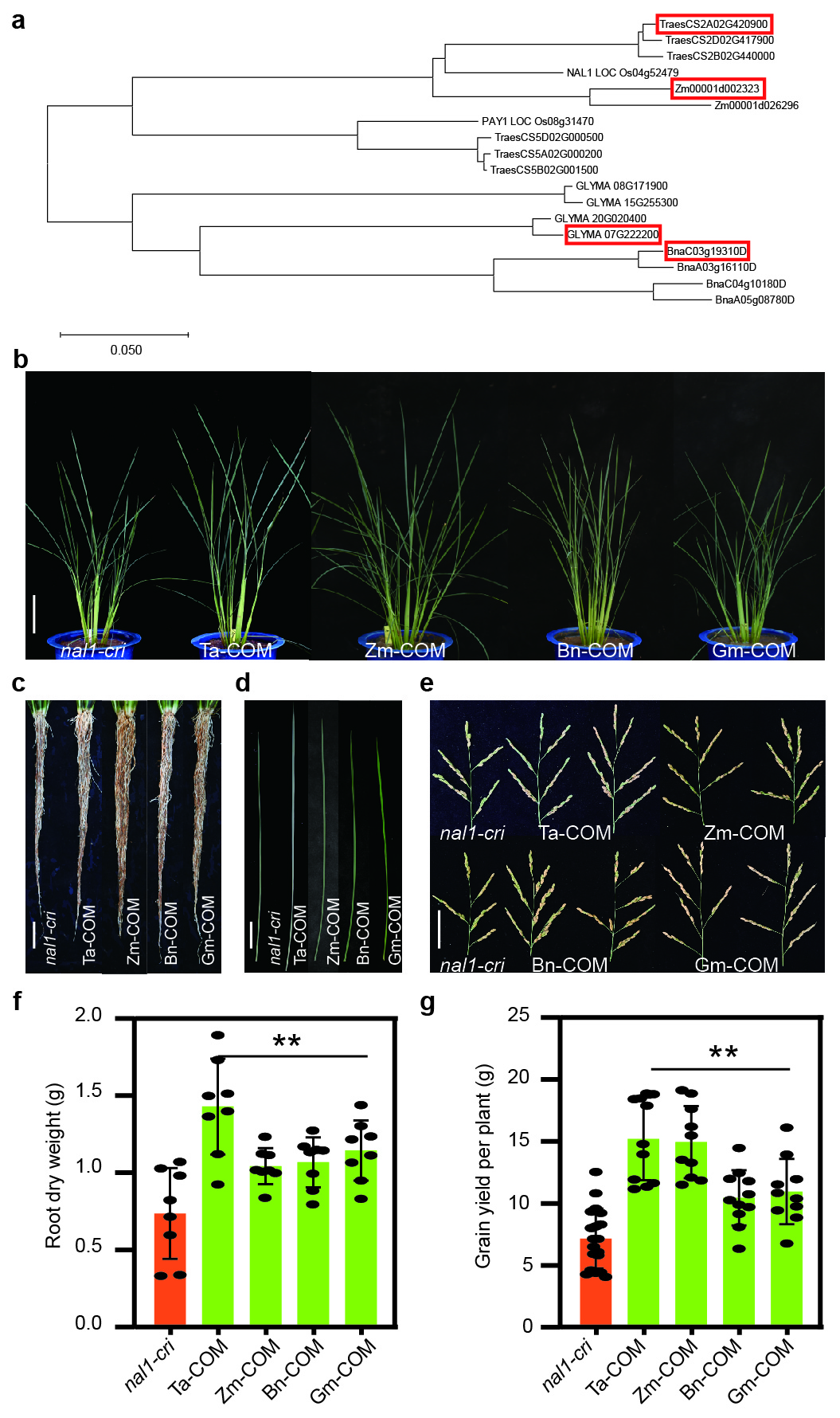
Since NAL1 is a breeding-valuable pleiotropic gene in rice, we further examined the conservativeness of the function of its homologs in other crops. A phylogenic tree of NAL1 and its homolog proteins from wheat, maize, soybean, and rapeseed showed that all the four crops had homologs of NAL1 (Extended Data Fig. 7a). One homolog from each crop was selected to complement nal1-cri. Compared to nal1-cri, the complementary lines exhibited significant recovery in plant height, flag leaf morphology, root system, and grain yield (Extended Data Fig. 7b-g), indicating that the NAL1 homologs from main crops may have conserved functions.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}