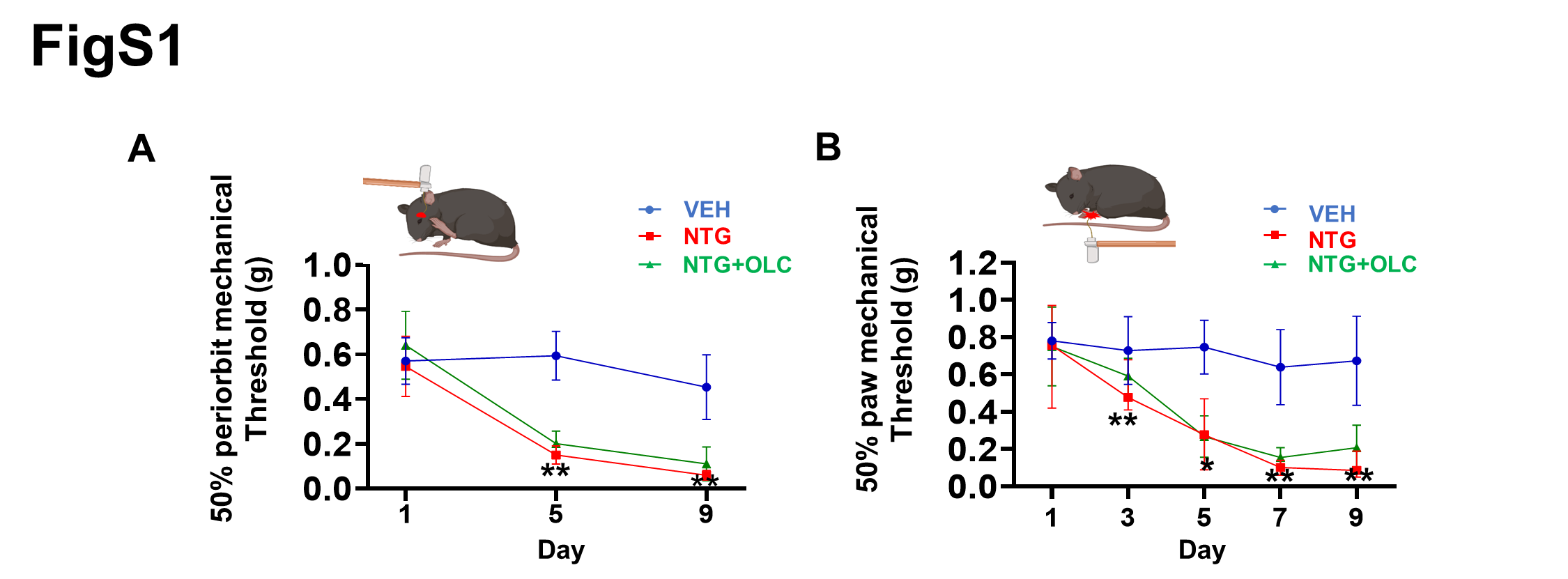
Herein, we investigated changes in brain activation with olcegepant treatment by assessing c-Fos expression in two chronic migraine models and found that the two chronic migraine models resulted in acute and chronic basal mechanical hyperalgesia, and the medial prefrontal cortex and caudal spinal trigeminal nucleus were both activated (Fig. 9A, B). Meanwhile, olcegepant alleviated hyperalgesia of the hind paw and periorbital area and activation of several brain regions. These changes in brain regions help us better understand the pathogenesis of chronic migraine, providing the foundation for improved treatment options and future research.
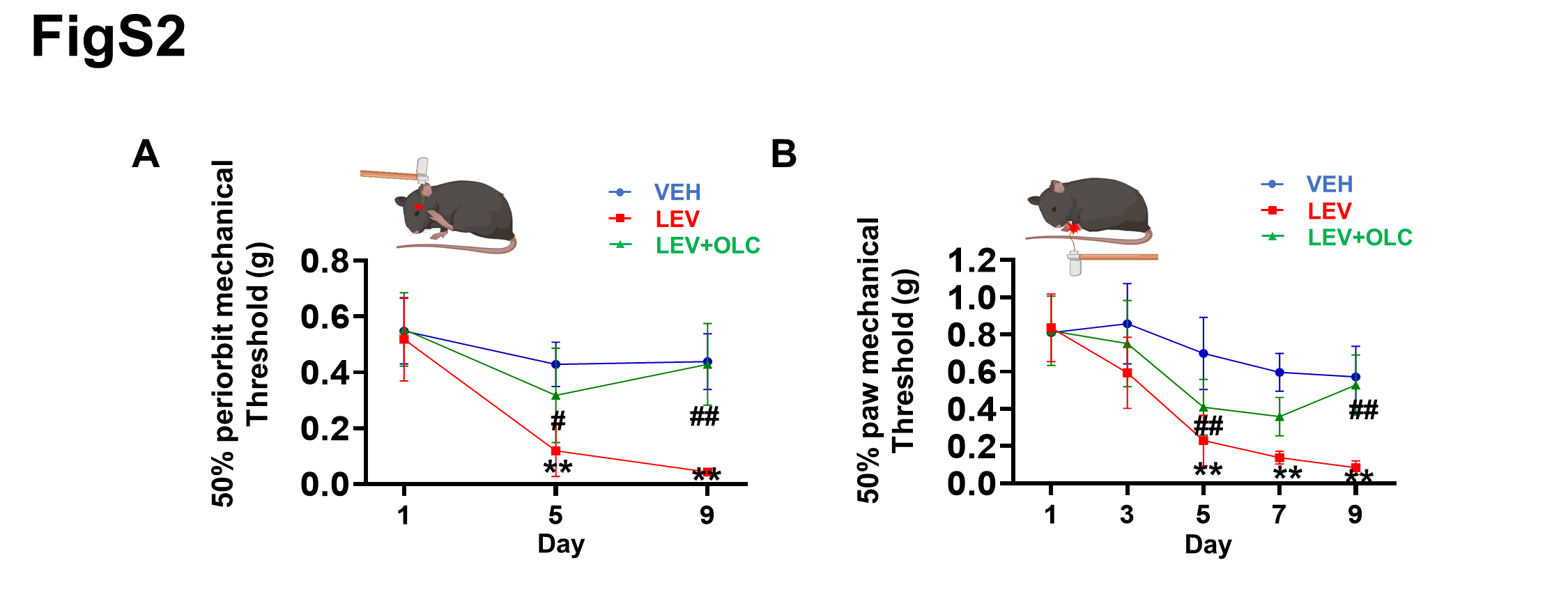
Olcegepant alleviated acute mechanical hyperalgesia and failed to prevent basal hyperalgesia in NTG-induced migraine mice. Recent studies have reported similar results explained by the relatively short plasma half-life of olcegepant in rodents [23, 26]. However, in LEV-induced chronic migraine mice, olcegepant alleviated both acute and basal hyperalgesia. Although the CGRP signaling pathway and functional smooth muscle KATP channels of extracerebral origin were shared in both chronic migraine mice models [23–24, 33], NTG can reportedly activate the sensory chemoreceptor channel TRPA1 of trigeminal afferents rather than LEV [23, 34]and increase BBB permeability and promote central neuroinflammation [35], explaining the difference in the treatment in both CM models. Meanwhile, LEV can cross the blood-brain barrier [36] and induces KATP channel opening leading to the triggering of aura and migraine headache through distinct mechanisms in humans [20, 21]. It is widely thought that LEV may increase the extracellular potassium concentration in glial cells, cortical neurons and cerebral vasculature [37] or lead to stimulation of hyperpolarization-activated cyclic nucleotide-gated cation channels (HCN channels) [38], resulting in the generation of cortical spreading depolarization (CSD), the most likely underlying mechanism of migraine aura [39]. The CGRP receptor is widely expressed in the central nervous system [40–43], and olcegepant can reduce CSD [44], relieving basal hyperalgesia by targeting central CGRP receptors in LEV-induced chronic migraine mice. However, LEV’s ability to induce CSD remains largely underinvestigated, warranting further research in animal models.
Similar c-Fos activity in the medial prefrontal cortex was found in both CM models, and olcegepant reversed brain activation, which may be due to the fact that CGRP signaling way is shared [23–24] between both chronic migraine mice models, and CGRP receptors are distributed in the cortex [41–42]. Meanwhile, in CM patients, resting-state functional connectivity of the default mode network (DMN) also decreased, including ROIs of the lateral parietal and medial prefrontal cortex [7] and headache frequency was negatively correlated with the volume of the medial frontal lobe cortex [12]. These studies also corroborated that activation of medial prefrontal cortex involves migraine chronification. In NTG-induced migraine mice, the activation of ACC was consistent with clinical imaging studies and the finding of other genetic mice migraine models. For instance, in CM patients, stronger structural connectivity was found between the caudal anterior cingulate cortex (ACC) and other brain regions [5, 10] and the N-acetyl-aspartate of bilateral thalami and right anterior cingulate decreased [6]. In the familial hemiplegic migraine type 2 (FHM2) mouse model, migraine-relevant hypersensitivity triggered by NTG has been attributed to altered neural function in the cingulate cortex [45] and CGRP receptors distributed in the cingulate cortex [39]. However, the ACC did not show high c-Fos activity in LEV-induced migraine mice. Unlike NTG, LEV can reportedly target functional smooth muscle KATP channels of extracerebral origin [33] or may induce CSD [23–24, 34–35], explaining the difference in brain activation in the two CM models.
Moreover, NTG-induced chronic migraine model with early growth response gene 1 (Egr1)-enhanced green fluorescent protein (EGFP) transgenic mice did not reveal any difference in the medial prefrontal cortex and no change in Egr1 expression was found in the anterior cingulate area (ACC) [18]. These results were also explained due to analysis of different immediate-early genes (IEGs) markers of active neurons.
Migraine chronification maybe involve thalamic contribution to central sensitization [4]. In this study, neuronal activity was increased in the PVT in LEV-induced migraine mice. It is well-established that PVT plays a critical role in the central processing of chronic pain [46]. However, no change in neuronal activity in the PVT was found in NTG-induced migraine mice. The PVT-vlPAG circuit reportedly mediates the central mechanisms of descending pain facilitation underlying persistent pain [47]. Although several clinical studies showed that vlPAG, the descending pain-modulating system, was associated with allodynia [15, 48], vlPAG did not exhibit high c-Fos activity in both CM models. In other migraine animal models, cortical spreading depression decreased c-Fos expression in rat PAG [48], and inflammatory soup administration increased c-Fos expression of PAG [49]. Thus, the role of vlPAG and PVT may differ in various chronic migraine models. Interestingly, Sp5c was activated in two CM models. CM patients also exhibited cephalic and extracephalic allodynia, corresponding to the sensitization of the second-order neurons in the caudal spinal trigeminal nucleus [50]. Consistent findings have been reported in the literature [28, 29]. Our results further substantiated that Sp5c is involved in migraine chronification. As expected, olcegepant alleviated the activation of Sp5c. Immunohistochemical studies performed with rat and human tissue revealed that the CGRP receptor was expressed in the medulla-pons region, cervical spinal cord [40, 43] and NTG induced an increase in the gene expression of CGRP and c-Fos in the spinal trigeminal nucleus. Our findings suggest that olcegepant may act on the CGRP receptor of the Sp5c to attenuate neuronal activity.
There were several strengths and limitations in this study. To the best of our knowledge, this is the first study to document the c-Fos activation map of pain relative to brain regions in NTG-induced migraine mice and LEV-induced migraine mice and provided a reference for neural circuit study in future research. However, we only focused on five representative coronal sections. These results may not necessarily be observed in CM patients, and the role of other brain regions cannot be completely excluded. Besides, since olcegepant alleviated acute hyperalgesia but not basal hyperalgesia in the NTG mice model, mice were sacrificed two hours after NTG or LEV injection on day 9. A recent study showed that basal hyperalgesia was sustained in NTG-induced migraine mice 24 hours after the last NTG injection [30]. Our future studies will focus on understanding whether these brain regions are still activated.
{kind=link}
{kind=link}