FABP5 is downregulated in CRC
To establish the precise function of FABP5 in cancer, we investigated its expression patterns in various tumor types. FABP5 was overexpressed in renal clear cell carcinoma and liver cancer and, conversely, downregulated in lung, breast and colon cancer (Supplementary Fig. 1A). As few reports have explored the function of FABP5 in CRC, we initially established a tumor microarray using 90 CRC and peri-tumorous tissues. Consistent with findings from the CPATC database, our results showed downregulation of FABP5 in CRC (Fig. 1A–D). Moreover, the FABP5 mRNA level was lower, but not to a significant extent, in tumor than control tissues (Supplementary Fig. 1B). Data from survival analyses indicated better prognosis of patients with higher FABP5 levels (Supplementary Fig. 1C).
Overexpression of FABP5 is associated with reduced CRC cell proliferation, migration and invasion
To ascertain whether FABP5 plays a tumor suppressor role in CRC, stable FABP5-overexpressing HCT116 and SW620 cell lines were generated via stable transfection. Western blot results validated the efficiency of FABP5 overexpression (Fig. 2A). In CCK-8 and EdU assays, upregulation of FABP5 was concomitant with suppression of cell proliferation (Fig. 2A–C; 2F–G). Moreover, data from the clone formation assay showed decreased clone numbers in FABP5-expressing HCT116 and SW620 cells (Fig. 2D–E). In the transwell assay, upregulation of FABP5 suppressed cell migration and invasion (Fig. 2H-J). Overall, FABP5 functioned as anti-cancer in CRC.
Knockdown of FABP5 promotes CRC cell proliferation, migration and invasion
To further establish the anti-cancer function of FABP5 in CRC, HCT116 and SW620 cell lines with stable knockdown of FABP5 were generated. Western blot analysis validated the efficiency of FABP5 knockdown (Fig. 3A). In CCK-8 and EdU experiments, downregulation of FABP5 promoted cell proliferation (Figs. 3A–C, 3F–G). Notably, suppression of FABP5 increased the clone numbers of HCT116 and SW620 in the clone formation assay (Fig. 3D–E). Data from the transwell assay showed that FABP5 silencing facilitated migration and invasion of HCT116 and SW620 cells (Fig. 3H–J), supporting a tumor suppressor role of FABP5 in CRC cells.
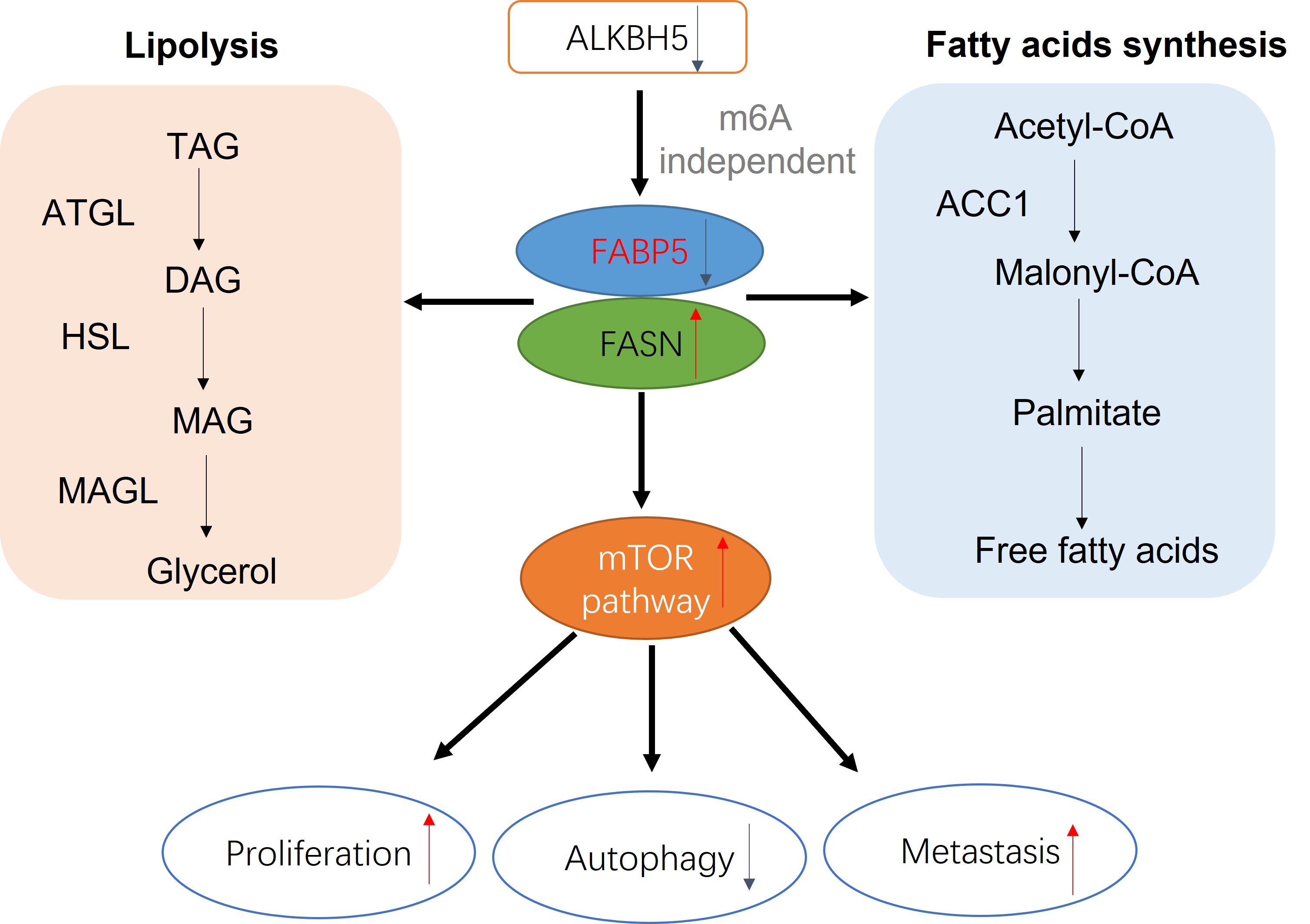
FABP5 interacts with FASN and promotes its ubiquitin proteasome pathway
With the aid of combined immunoprecipitation and mass spectrometry analyses, 395 proteins interacting with FABP5 were identified. Among these proteins, FASN attracted us attention, which was a fatty acid synthase. Subsequent co-IP experiments confirmed interactions of FABP5 with FASN (Fig. 4A). Notably, knockdown of FABP5 led to an increase in FASN expression and, conversely, overexpression of FABP5 induced a decrease in FASN (Fig. 4B). After treatment with CHX, the stability of FASN was increased in FABP5 knockdown and decreased in FABP5 overexpression groups. Moreover, the FASN level was markedly increased upon MG132 treatment (Fig. 4C). Co-IP assay of FASN and ubiquitin consistently showed that knockdown of FABP5 led to a decrease in ubiquitin combined with FASN while overexpression of FABP5 enhanced this binding (Fig. 4D). To further resolve its function in CRC, FASN was inhibited via shRNA or treatment with orlistat. Under conditions of knockdown of FASN (Fig. 4E, Supplementary Fig. 2A), cell proliferation (Fig. 4F, H; Supplementary Fig. 2B, 2E-F), clone formation (Fig. 4G, Supplementary Fig. 2C-D), cell migration and invasion (Fig. 4I; Supplementary Fig. 2G-H) were inhibited. Similar phenomena were observed in the orlistat treatment group (Fig. 4J-M; Supplementary Fig. 2I-Q). The collective results clearly indicate that FABP5 regulates FASN via stimulation of its ubiquitin proteasome pathway.
Knockdown or inhibition of FASN suppresses malignant biological behaviors activated by downregulation of FABP5
Downregulation of FABP5 promotes malignant biological behavior in CRC. To explore whether oncogenic activity is mediated by upregulation of FASN, knockdown of FASN or orlistat treatment in FABP5 downregulation stably transfected cells was performed. Firstly, efficiency of knockdown was detected via western blot (Fig. 5A; Supplementary Fig. 3A). CCK-8, colony formation and EdU experiments revealed that downregulation of FASN or inhibition of its activity led to suppression of cell proliferation promoted by knockdown of FABP5 (Fig. 5B-F; Supplementary Fig. 3B-F). In the Transwell assay, silencing or inhibition of FASN reversed the increase in migration and invasion induced by downregulation of FABP5 (Fig. 5G, H; Supplementary Fig. 3G, H). Nile red staining demonstrated that overexpression of FABP5 decreased while its knockdown increased lipid accumulation (Fig. 5I). FASN depletion resulted in attenuation of lipid accumulation induced by FABP5 silencing, highlighting a key role of FASN in FABP5-regulated malignant behavior(Fig. 5J). Moreover, sole knockdown of FASN in wild-type CRC consistently led to a decrease in lipid accumulation (Fig. 5K). These results suggest that oncogenic progression initiated by downregulation of FABP5 is restored by FASN inhibition in CRC cells.
FABP5 promotes autophagy via inactivation of the mTOR pathway mediated by FASN
To explore the mechanisms underlying the tumor suppressor role of FABP5 in CRC, RNA -seq and lipid-omics were performed in FABP5 overexpression and FASN knockdown along with the respective control groups. Multiple genes associated with numerous biological processes and signaling pathways were dysregulated, including TGF-β, Hippo, Wnt, NF-κB and mTOR, as observed via RNA-seq (Fig. 6A, C; Supplementary Fig. 4A. Lipid-omic analyses further revealed that FABP5 and FASN regulate autophagy through effects on lipid metabolism (Fig. 6B, D; Supplementary Fig. 4B). Western blot results confirmed that upregulation of FABP5 promotes while knockdown of FABP5 inhibits autophagy (Fig. 6E). Moreover, silence of FASN or treatment with orlistat rescued inhibition of autophagy induced by FABP5 silencing (Fig. 6F, Supplementary Fig. 3G). Similar to this finding, overexpression of FABP5 suppressed, while knockdown of FABP5 activated, the mTOR pathway (Fig. 6G). Both silencing of FASN and treatment with FASN inhibitor inactivated the mTOR pathway (Fig. 6H; Supplementary Fig. 3I; Supplementary Fig. 4C-F), supporting a tumor suppressor role of FABP5 via inhibition of mTOR through FASN.
Overexpression of FABP5 and knockdown of FASN inhibits tumor growth in vivo
We further examined the roles of FABP5 and FASN in vivo with the aid of tumor xenograft models. To this end, SW620 cells transfected with FABP5 overexpression or FASN knockdown vector, FABP5 knockdown vector plus orlistat and control vector were implanted in nude mice. Notably, the weights and volumes of tumors were decreased in mouse xenografts injected with SW620 cells bearing FABP5 overexpression and FASN knockdown vectors (Fig. 7A–F). Conversely, SW620 cells with stable FABP5 knockdown showed accelerated tumor growth compared with the control group, which was suppressed by orlistat (Fig. 7G–I). Immunohistochemical staining of FABP5 and FASN revealed FABP5 negatively regulated FASN expression in vivo (Fig. 7J-L). Our collective results support an essential role of FABP5 in FASN-mediated CRC progression, both in vitro and in vivo.
ALKBH5 regulates FABP5 in an m6A-independent manner
Next, we focused on the mechanisms underlying downregulation of FABP5 in CRC. The m6A prediction server, SRAMP, revealed abundant m6A modification sites in FABP5. Upon knockdown of m6A writers and erasers (METTL3, METTL14, WTAP, FTO and ALKBH5), positive regulation of FABP5 was observed in the absence of ALKBH5 (Fig. 8A, B). Furthermore, ALKBH5 negatively modulated the FASN level (Fig. 8C–E). To explore whether ALKBH5 inhibited CRC via FABP5, we additionally depleted FABP5 in ALKBH5-overexpressing cells (Fig. 8F). The anti-cancer effects of ALKBH5 (including suppression of cell proliferation, migration and invasion) were partly restored by silencing of FABP5 which indicated FABP5 could be regulated by ALKBH5(Fig. 8G–M).
{kind=link}