White Spot Syndrome Virus (WSSV) continues to be the most severe viral pathogen to the shrimp industry worldwide. Pacific white shrimp Litopenaeus vannamei is particularly affected by WSSV and this virus has been detected in a wide range of wild crustaceans, including penaeid and non-penaeid shrimp, crayfish, as well as crabs. Crabs have been considered as a dangerous threat to shrimp farms because they are generally believed to be highly tolerant to WSSV and to remain infected for long periods without signs of disease. The burrowing crab Neohelice (= Chasmagnathus) granulata can be commonly found in the surroundings of the shrimp farms in southern Brazil. Here, we investigated the susceptibility of N. granulata to WSSV infection in comparison to L. vannamei. WSSV infectability and host susceptibility were investigated by viral challenge (intramuscular injection) in both species. Viral load in challenged animals was quantified by qPCR in both hemolymph and gills. Furthermore, the transcript levels of sixteen target genes related to the molecular defense status were assessed. The results revealed that N. granulate experimentally infected by WSSV, as well as those naturally infected, showed lethargy, lack of appetite and later gross signs of the disease. Moreover, N. granulate showed to be less susceptible to WSSV when compared to L. vannamei. While no death was observed in crabs before a post-challenge interval of 120 h; shrimp death was seen as early as 48 hours post-infection. Comparative viral load was also assessed by qPCR in gills of captured wild crabs and farmed shrimp naturally infected by WSSV. Transcript levels of target genes were also investigated after WS-challenge between 24 and 48 hpi in shrimp and between 96 and 120 hpi in crab. Differences in gene transcripts were particularly noteworthy with the increase of viral copies at 106 copies.µl− 1. These results indicated that WSSV infection modulated genes related of important cellular responses, such as apoptosis, chaperone function and ion storage, in both shrimp and crab.
Research Article
Susceptibility of Neohelice granulata (Decapoda, Varunidae) to White Spot Syndrome Virus (WSSV)
https://doi.org/10.21203/rs.3.rs-1971357/v1
This work is licensed under a CC BY 4.0 License
You are reading this latest preprint version
Neohelice granulata
Litopenaues vannamei
WSSV
Viral load
qPCR.
Viruses are extremely abundant in aquatic systems (Suttler, 2005, Zhang and Gui, 2018) and are considered as the most important pathogens in impacting shrimp farming worldwide. Different life stages of shrimp may be susceptible to viral infections which can cause mortality, slow growth and deformations (Rahman et al., 2007, OIE, 2016). Among the several viruses that have been reported as pathogenic to shrimp, White Spot Syndrome Virus (WSSV) continues to be one of the most severe and associated to huge global economic losses to the shrimp farming. Since the first report of WSSV in Brazil, in the south of Santa Catarina state in January 2005, the disease was soon detected in some farms located in the northern and central regions of the state, as well. Subsequently, the southern region of the state was isolated, and the stocking suspended until September of that year (Seiffert et al., 2005). After the fallowing, new stockings took place, but successive losses were still reported in subsequent years. Since then, WSSV outbreaks have been seen in many areas of Santa Catarina, as well as in other regions of Brazil until now. WSSV caused a drop of more than 90% in the production of shrimp in Santa Catarina (Goularti and Ronçani, 2018).
WSSV is a double-stranded rod-shaped DNA virus, which belongs to Nimaviridae family (Mayo, 2002), being unique among shrimp viruses due to its large genome size of approximately 290 kbp (van Hulten et al., 2001). Clinical signs of White Spot Disease (WSD) caused by WSSV include lethargy, reddish discoloration of the animal’s body, and emaciation of the carapace, which may display the presence of white spots (Durand et al., 1997). Mortalities on cultured shrimp may reach 100% within 3–10 days after exhibiting the first clinical signs of the disease. Survivor shrimp may carry the virus for life and may also pass the virus to their progeny (Lo et al., 1996a, 1997a).
Water discharges from shrimp farms were seen as the source for wild crustacean population infection (Lo et al., 1996a, b), and the consequent high prevalence of WSSV in natural populations (Meng et al., 2009). WSSV propagates quickly throughout the environment and has been found not only in wild penaeid shrimp but also in a wide host range, including crab, lobster, copepod, crayfish and freshwater crab and prawn, as well (Lo et al., 1996a, b; Maeda et al., 1998; Peng et al., 1998; Wang et al., 1998; Sahul Hameed et al., 2001, 2003; Marques et al., 2011; Sánches-Paz et al., 2015). Mortality among this host range is highly variable according to several reports, suggesting that susceptibility varies between crustacean taxa (Oidtmann and Stentiford, 2011). Economic impacts have driven research on WSD in farmed shrimp, especially in Litopenaeus vannamei, and reports in the literature are abundant for this penaeid species. In contrast, the data concerning the effects of the disease in wild noncommercial crustacean populations remain scarce.
A wild crustacean frequently seen in the surroundings of shrimp farms in south of Brazil is the crab Neohelice granulata (Dana, 1851), previously known as Chasmagnathus granulate, before Sakai et al. (2006) revised and reclassified all species formerly attributed to the genera Helice and Chasmagnathus. N. granulata is a burrowing semi-terrestrial crab found in the intertidal zone of estuaries, salt marshes and mangroves of the South-western Atlantic Ocean, ranging from San José Gulf, northern Patagonia, Argentina, throughout Uruguay, to Lagoa Araruama, in Rio de Janeiro, Brazil (Spivak, 2010). In southern Santa Catarina, it can be found along the extension of the estuary of the huge Laguna Lagoon Complex, around which most of the shrimp farms are located. This crustacean is abundant in the rearing ponds, digging holes and galleries on the margins (Marques et al., 2011).
N. granulata has been previously reported as a carrier and as a potential host of WSSV, due to natural infection, based on sequence homology data to the viral genome of a nested PCR fragment obtained from specimens collected in Santa Catarina, Brazil (Marques et al., 2011). Later, positive PCR results were also reported for N. granulata collected further south, in Rio Grande do Sul, Brazil (Cavalli et al., 2013).
Crabs have been considered as a particularly dangerous threat to shrimp farms because they are generally believed to be highly tolerant to WSSV and to remain infected for long periods of time without showing signs of the disease, therefore contributing to the maintenance of the viral agent in the environment. Since N. granulata susceptibility to WSSV has not been addressed so far, our aim was to carry an investigation under a comparative perspective to L. vannamei, including the analysis of the transcript levels of target genes related to molecular defense responses.
The identification of potential hosts in penaeid farms, as well as surrounding areas, may contribute to monitoring the presence and behavior of the virus in the environment, as well as to determine its geographic distribution, and provide information about its persistence and pathogenicity (Marques et al., 2011; Moser et al., 2012). These aspects should be considered as a critical topic in sanitary programs and to reinforce management strategies.
2.1. Animals
Crabs, Neohelice granulata (n = 115; 5.3 ± 0.6 cm in body length; weight 12.7 ± 1.1 g), were obtained nearby the tanks of a local shrimp farm in Northern of Santa Catarina State (Brazil), transported to the laboratory, and maintained in 30 L fiberglass tanks (n = 4), containing a thin layer of filtered sea water, at 23 ± 1°C and salinity of 25 g.L− 1. Crabs were held under these conditions for 5 days to acclimate prior to viral challenge. During this period, crabs were fed once a day with commercial shrimp feed. Crab identification was carried out according to the key to Brachyura of the Brazilian coast (Melo, 1996).
Juvenile intermolt shrimp L. vannamei (n = 110; 8.5 ± 2 cm in body length, weight 10 ± 1 g) were obtained from the same shrimp farm, transported to the laboratory and maintained in 6L fiberglass tanks (n = 3), with filtered sea water at 23 ± 1°C and salinity of 20 g.L− 1. Shrimp were held under these conditions for 5 days to acclimate prior to viral challenge. During this period, shrimp were fed once a day with commercial shrimp feed.
To investigate the previous presence of Infectious Hematopoietic Necrosis Virus Hypodermal (IHHNV) and White Spot Syndrome Virus (WSSV), hemolymph samples were used for PCR assays. After IHHNV and WSSV screening, only animals that showed to be negative for both viruses were used for the challenge experiments.
Additionally, to better access the natural WSSV prevalence, a total of other 30 N. granulata crabs (5.8 ± 1 cm in body length; weigh 12.5 ± 5 g) and 30 L. vannamei shrimp (14.5 ± 3 cm in body length; weigh 20 ± 2 g) were collected in another shrimp farm, located in the southern part of the state. In the case of these other specimens, gill samples were used to WSSV screening.
2.2. Viral challenge
Crabs and shrimp were challenged with WSSV by injection, using an inoculum corresponding to a Brazilian geographical isolate of the virus, previously characterized (Müller et al., 2010). The inoculum was prepared as follow: WSSV-infected L. vannamei tissues were diluted 1:5 (w/v) with sterile PBS buffer (pH 7.8) and homogenized. The homogenate was clarified by centrifugation, filtered (0.45 µm filter) and used for injection. Prior to injection, the concentration of WSSV inoculum was assessed by qPCR. A standard curve was used for viral quantification, with serial dilutions of recombinant plasmid containing a previously known number of WSSV genome sequence copies per µL. First crabs (n = 56) were injected with 10 µL purified WSSV (5.3x104 viral copies per animal) via the base of the walking leg using a sterile 1 mL syringe fitted with a 21G needle. The control group was injected with the same volume (10 µL) of PBS buffer. Subsequently, shrimp (n = 54) were injected with 100µL purified WSSV (5.5x104 viral copies per animal) intramuscularly into the third dorsal segment. Likewise, the same volume of PBS buffer was used for injecting the control group. Crab experiments were performed in duplicate, whereas shrimp experiments were performed in triplicate. After challenge, crabs and shrimp were monitored daily for survival, pattern of behavior, mobility, and body coloration, as well as feed consumption.
2.3. Sampling and DNA extraction
In the challenge experiments, gill samples were used to monitor viral infection at intervals of every 24-hour post-infection (hpi). Samples were collected from eight crabs and nine shrimp at each time. The first sampling was performed prior to infection (0 hour) and the last one took place at 72 and 144 post-infection (hpi), for shrimp and crab, respectively. Accordingly, gills from natural infected farmed shrimp and wild crabs were also used to assess viral infection. DNA extraction from gills was performed according to Marques et al. (2011). The concentration of total DNA of gills was calculated by the optical density (OD) ratio at 260/280 nm using a NanoDrop® 2000 spectrophotometer reader (ThermoScientific).
2.4. WSSV detection and viral load determination
A standard conventional polymerase chain reaction (PCR) was used for initially screen for WSSV natural infection in N. granulata and L. vannamei samples, using primers designed to amplify a 480bp fragment of the WSSV genome, according to Nunan and Lightner (2011). PCR products were submitted to 2% agar electrophoresis and qualitative results were documented by digital camera. Viral load determination in these samples, as well as for both WS-challenged N. granulata and WS-challenged L. vannamei was performed by qPCR. An amount of 100 ng.µl− 1 DNA was added to a reaction mix prepared according to the protocol of QuantiTect® SYBR® Green PCR-Kit (Qiagen, Hilden). Reaction was performed in a thermocycler (Rotor-Gene Q), with an initial denaturation at 95°C for 15 min, followed by 39 cycles of 95°C for 30 sec, annealing temperature at 55°C for 30 sec, and extension at 72°C for 10 sec. The set of primers were designed based in the sequence amplified by the nested PCR reaction described by Lo et al. (1996a): WS2F: 5’-TGCCTTGCCGGAAATTAGTGTGTG-3’ and WS2R: 5’ACAACATCCAACAATGGTCCCGTG-3´. After amplification resulting were analyzed and compared to the standard curves. Standard curves were performed with a serial diluted DNA from a recombinant plasmid containing a previously known number of WSSV genome, belonging to our laboratory collection, as internal positive controls.
Table 1. Primers used in the analysis of differential expression by RT-qPCR in Neohelice granulata and Litopenaeus vannamei.
|
Gene |
Functional category |
Primer sequence (5’ – 3’) |
Code |
|---|---|---|---|
|
Calreticulin |
Calcium homeostasis |
F- CTCTTTCGCTCCTCTTCATC R- CCTCATCTCCAATGATCCTG |
CALR |
|
Calcified cuticle protein |
Crustacean cuticle formation |
F- GACAGCAGTCCTGTAGACAAGAAG R- GTTAGGGAAAGTTGGACTCAC |
CALC |
|
Ferritin |
Iron storage protein |
F- ATCCAGAAGACGATCAAG R- GTTCTCCAGCAGATTGCAG |
FERR |
|
Caspase 2 |
Apoptotic process |
F- TAATCCAAGACCCGGAACTGGCAA R- CAACCTTCCATTCATTCCGTGCGT |
CASP-2 |
|
theCaspase 3 |
Apoptotic process |
F- TTGTTTCTGGCTTTGACTTCGCCG R- GAGTGTCGAGTGGATGTAAG |
CASP-3 |
|
ProPhenoloxidase |
Prophenoloxidase system |
F- TGAAAAAGTGGGTCGAGGAC R- AACACTGAGTCTTTCCATGTCGT |
ProPO |
|
Phenoloxidase |
Prophenoloxidase system |
F- CGGTGACAAAGTTCCTCTTC R-GCAGGTCGCCGTAGTAAG |
PHEN |
|
QM Protein |
Apoptotic protein |
F- GATAACGTGGTAAGGGTGTAGC R- CGTATCTGCGCCAACAAGTATC |
QM |
|
Inhibitor of apoptosis protein |
Apoptotic process |
F- GTTACTTCACCACAGTCCACTAA R- CATCCATCTCGGCATCCATAC |
IAP |
|
Ubiquitin |
Protein modification process |
F- AAGGCCAAGATCCAGGATAAGGAG R- AGTCAGACAGAGTGCGACCATCTT |
UBC |
|
Heat Shock Protein-60 |
Protein modification process |
F- GGCATCCACAGCCAACATAA R- TGCCAACAACACCAACGA |
HSP60 |
|
Heat Shock Protein-70 |
Protein modification process |
F- ACAGACACAGAGCGTCTGATTGGT R- TTGATGATGGTGAAGGGCCAATGC |
HSP70 |
|
Protein Kinase C |
Intracellular signaling cascade |
F- CTGTCCTGTGTAAGGTTCTC R- CCATAGTGGTTGGTCTTGAG |
PKC |
|
Histone2ɑ |
Nucleosome formation |
F- AGTACTTGGCTGCTGAAGTC R-CTCTTCGTCGTTACGGATGG |
H2ɑ |
|
β-actin |
Cytoskeleton protein |
F- AGCCTTTCGCCTGAACATAGCAGT R- TGATCACCATCGGCAACGAGAGGTT |
β-ACT |
|
β-tubulin |
Cytoskeleton protein |
F- AAGATGGCCTCTACCTTCATCGGT R- AGCCTTTCGCCTGAACATAGCAGT |
β-TUB |
|
18S Ribosomal (housekeeping gene) |
F- GGCCTCGCAATTGGAATGAGTACA R- TACGAGCGTTTCAACCGCAAC |
DEC |
2.5 Transcript levels of target genes in samples of N. granulata and L. vannamei infected with WSSV.
Gills collected from WSSV challenged animals and their respective controls were used to investigate differential transcription of target genes. Total RNA from the gill samples was extracted using the TRIzol reagent (Invitrogen), according to the manufacturer's protocol. Total RNA concentration and purity was assessed by spectrophotometer (NanoDrop 2000, ThermoScientific) through the absorbance at 260 nm and 260 nm/280 nm ratio, respectively. Total RNA from crab and shrimp gills was reversely transcribed using Quantitect Reverse Transcription Kit (Qiagen, Hilden). Diluted cDNA (100 ng.µl− 1) was added to a reaction mix composed of QuantiFast®SYBR®Green PCR kit (Qiagen, Hilden). Amplifications were performed in duplicates for each sample using a Rotor-Gene 6000 real-time PCR thermocycler (Qiagen, Hilden) under the following conditions: 1 cycle at 95oC for 5 min; 30–40 cycles of amplification at 95oC for 10 s, 58–60oC for 30 s. A dissociation curve was carried out to confirm the amplification of a single product. Efficiency of PCR reaction was determined using a standard serial dilution curve of cDNA and negative control. Transcript levels of sixteen target genes were quantified by the method of 2− CT, using Ribosomal 18S-like as a house-keeping normalizing gene (Schmittgen and Livak, 2008) and the Rotor-Gene 6000real-time qPCR system software. The analyses were performed using GraphPad Prism 5.0 software (GraphPad Software). The primer designing was based on our previous studies assessing shrimp defense responses through gene transcription and proteomics (Müller, 2009; Valentim et al., 2014, 2015; de Souza Valente et al., 2020; Mattos et al in preparation) (Table 1).
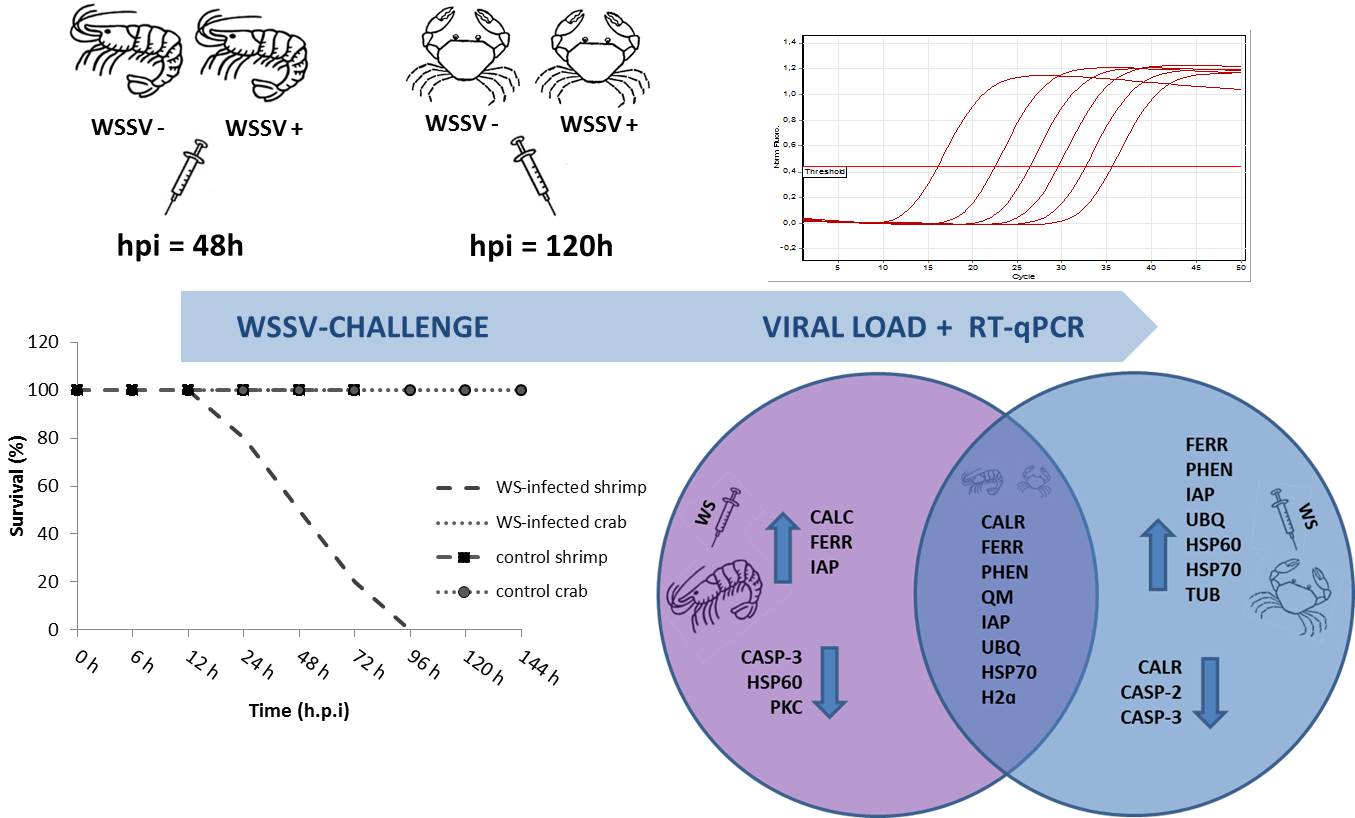
Viral challenge with injection of WSSV resulted in infection of burrowing crab N. granulata. All WSSV-challenged crab showed PCR positive reactions, evidencing the presence of the virus. WSSV challenge N. granulata survived up to 168 hours, with no gross signs of infection. Only on the sixth day (144 hours) post-infection it was possible to observe lethargy and reduction in food consumption when compared either with earlier days or with control group. On the other hand, WSSV challenged shrimp showed lethargy, reddish body color and low mobility, clinical signs associated to WSSV, earlier as 48 hpi. Mortalities were recorded at 48 hpi. At 72 hpi all shrimp were collected. Comparative results show wide variation in responses to WSSV challenge. Although both, N. granulata and L. vannamei, were susceptible to WSSV, L. vannamei showed to be more susceptible than N. granulata. Crab and shrimp survival curves after WSSV-challenge can be seen in Fig. 1.
3.1. Real-time PCR detection and viral load after WS challenge
Gills were used for WSSV detection and determination of viral load, by real-time PCR (qPCR). qPCR showed that WSSV could replicate within the crab tissues, as well as in shrimp (Fig. 2). Through these samples, it was possible to evaluate the progress of WS infection in challenged animals. Results showed WSSV-positive crabs in the challenged group, as expected, while the control group remained negative for the virus during the whole experiment time course. Likewise, all shrimp, sampled after virus injection, were WSSV-positive, whereas the control group remained negative.
qPCR analysis revealed significant differences (p < 0.05) in the viral load of WS-positive crabs, according to the time interval following virus infection (Fig. 2A). After 24h, an amount of 3.59 copies were detected, which rose up to 2.14x105 (96 hpi) and further to 4.48x106 (144 hpi). Accordingly, increases in viral copies in WSSV-challenged shrimp were also seen (Fig. 2A). Nevertheless, after 24h the average number of viral copies in WSSV challenged shrimp was 8.38x101 copies.µl− 1, whereas after two days (48hpi), the average number of viral copies rose to 2.57 x106 and significantly decreased to 7.79 x105 copies at 72 hpi. qPCR average numbers showed that viral load in gill of shrimp at 48 hpi (2.57x106 copies.µl− 1) had a similar counterpart in crabs only at 120 hpi (4.39x106 copies.µl− 1). Also, it was possible to identify viral peaks at some post-WS infection phases. Comparatively, viral load has increased from 48 to 72 hpi in shrimp, while highest replication was detected in 96 to 120 hpi in crab (Fig. 2B). No viral copies were detected in control crab or control shrimp indicating virus-free animals.
3.2. WSSV PCR and qPCR detection on natural infected crustaceans
Thirty N. granulata crabs and thirty L. vannamei shrimp gills were also used to determine the viral load in naturally infected animals. The initial analyses were conducted by conventional PCR and PCR products visualized by electrophoresis in 2% agarose gel. PCR products that showed the presence of a single band equivalent to the expected molecular weight of approximately 480 bp were considered WSSV-positive animals. This band corresponds to the amplification of a specific fragment of the WSSV genome. The samples in which the 480 bp fragment was absent were considered negative for the presence of the virus. Furthermore, the viral load in WSSV positive field animals (crabs and shrimp) was measured by qPCR. In naturally infected field shrimps viral quantification (106 copies.µl− 1) was very close to those found in animals collected at 48 hours after experimental infection. The crabs, collected within or adjacent to shrimp farms, exhibited a lower amount of WSSV, ranging between 102 and 103 copies.µl− 1.
3.4. Differential gene transcription in WSSV challenged N. granulata and L. vannamei
Differential transcript levels in N. granulata and L. vannamei challenged with WSSV were assessed in gill samples by RT-qPCR, focusing on sixteen target genes. Comparison was made between two different post-infection time intervals (hpi) and their respective controls (Fig. 3). These data are summarized in Table 2 and show transcript levels at 24 and 48h post-infection in shrimp, and at 96 and 120h post-infection in crab.
Relative gene transcription profile to most of the selected genes showed significant statistical differences when compared to the respective controls. These findings reflect genes encoding essential molecular defense processes in both species. In crab, changes in transcript levels of fifteen genes were seen at 96 or 120 hpi, whereas in shrimp twelve genes showed significant changes at 24 or 48 hpi when compared with non-infected controls. Such genes showed an increase in expression in the range from 1.8 to 24-fold relative to control groups (Fig. 3).
According to qPCR results, experimentally infected shrimp displayed a total of viral particles around 106 after 48h of experimental infection (hpi). This value matches the viral load found in naturally infected L. vannamei collected in the field. On the other hand, the same amount of viral particles (106 copies.µl− 1) in N. granulata gills was seen only after 120h of experimental infection (hpi). Thus, samples containing the same amounts of viral particles were chosen to compare the induction or inhibition of some of the target genes. The higher viral load was associated to nine up-regulated genes in shrimp (CALC, FERR, CASP-2, PHEN, QM, IAP, UBQ, HSP70 and H2ɑ). This same viral load was associated to fourteen up-regulated genes in crab (CALR, CALC, FERR, CASP-3, PROPO, PHEN, QM, IAP, UBQ, HSP60, HSP70, PKC, H2ɑ and b-TUB). Moreover, when overall expression profile was analyzed and compared between species, out of the sixteen selected target genes, we found eight genes that were modulated by the severe infection. Significant changes were seen on transcript levels of CALR, FERR, PHEN, QM, IAP, UBQ, HSP 70 and H2ɑ in both species at similar viral load (106 copies.µl− 1).
Also, comparative analyses were made between WSSV-challenge groups, considering transcript levels and post-infection time intervals. In shrimp, upon viral load increase from 8.38 x101 to 2.57x106 copies.µl− 1 (24 to 48 hpi), CALR, CALC and IAP were induced, whereas CASP-3, HSP 60 and PKC were repressed. In crabs, FERR, PHEN, IAP, UBQ, HSP 60, HSP 70 and β-TUB were up-regulated while CALR, CASP-2, CASP-3 were down-regulated when viral load increased from 2.14 x105 to 4.39x106 copies.µl− 1 (96 to 120 hpi). Overall, these genes showed significant changes in transcript profile in response to the course of the infection and increase in viral load.
No significant changes were seen in the transcription of the genes encoding Prophenoloxidase or ß-Tubulin in shrimp after WS-challenge. Moreover, ß-Actin transcript levels showed no significant changes in shrimp or crabs.
Table 2. Differential expression in Neohelice granulata and Litopenaeus vannamei challenged with WSSV. Columns represent changes in transcript levels of sixteen target genes in shrimp at 24- and 48-hours post-infection (hpi), and in crab at 96 and 120 hpi. Average data represent differences in folds relative to respective control (blue and red bars). Mean statistical differences between WS-infected and non-infected animals (controls): (↑) up-regulated or (↓) down-regulated genes in time course.

4.1. Viral challenge and WSSV viral load
Many studies have investigated the presence of viral agents in the aquatic environment, as well as in cultivation systems, where crustaceans and other aquatic organisms may act as vectors (Flegel, 2006, Mijangos-Alquisires et al., 2006). However, relative few studies have considered the possibility of infection of wild animals generated by aquaculture activity itself. In this context, crabs Neohelice granulata collected in the vicinity of shrimp ponds were analyzed for the presence of WSSV and, in case of a positive result, had their viral load determined by quantitative real time PCR, as well. The viral load range fell between 102 to 103 copies.µl− 1. According to Shekhar et al. (2006), Souza (2008), and Walker et al. (2011), the degree of the viral infection severity can be predictable according to the viral load measured in the animals. So, based on the criteria pointed out in their work, wild crabs naturally infected by WSSV showed a light WSSV infection. On the other hand, qPCR results showed that naturally infected shrimp displayed 106 copies.µl− 1, corresponding to a severe WSSV infection.
Regarding the WSSV experimental infection, results showed that the adopted protocol was equally efficient for both crustacean species.
Animals challenged with the virus, showed positive reactions, when analyzed by nested PCR, indicating that the virus was present and, as a consequence, a potential infectious process was established. Moreover, qPCR analysis in previously nested PCR WS-positive individuals has revealed that WSSV was capable of replication within the tissues of shrimp, as expected, as well as in crabs. This is the first report of experimental infection in N. granulata crabs. Furthermore, to date, no work has shown the proliferation of the virus, based on the quantification of viral load through the time course of infection. An effective injection of WSSV into blue crab Callinectes sapidus has been reported by Blaylock et al. (2019) and in the mud crab Scylla serrate by Liu et al. (2011), both as examples of commercial species of interest for aquaculture. N. granulata investigated in the present study occurs widely throughout the state of Santa Catarina and is abundant in nurseries and cultivation pond areas, being part of the fauna accompanying and surrounding the farms (Marques et al., 2011). N. granulata has no commercial interest, unlike the blue crab. However, it can be considered as an emergent animal model for biochemical, physiological and ecological research (Spivak, 2010), besides comparative studies related to pathogenic infections in crustacea.
The qPCR results in gills of WSSV-challenged animals confirmed that the number of WSSV viral copies increased over time. Surprisingly, no clinical signs were seen in crabs along the progress of infection. Nonetheless, based on viral load results we can infer that the N. granulata crabs were effectively infected through laboratory viral challenge. Therefore, this crustacean specie serves as asymptomatic carriers of WSSV, and probably other viruses of high-prevalence, without developing the disease, which represents a potential source of reinfection of farmed shrimp and maintenance and spread of the pathogenic agent in the environment.
Viral particles present in water, sediment and vectors may represent important reservoirs and source of contamination, reintroducing diseases and making future crops unfeasible. According to OIE (2016) all crustacean species are potential hosts for WSSV. In addition, insects (Lo et al., 1996a), rotifers (Yan et al., 2004) and polychaetes (Vijayan et al., 2005) also function as vectors of the disease. These organisms can act as important vectors in the transmission of WSSV to shrimp, both in crops and in the wild. Thus, the presence of crabs in shrimp farms should be strongly avoided to disrupt this flow between infected and uninfected animals (Marques et al., 2011). An alternative to prevent the entry of larvae and juveniles of these vectors into nurseries is the use of 150–200 micron mesh (Balasubramanian et al., 2018). Chlorine water treatment also appears to be effective in eliminating vectors, such as crab larvae and copepods (Fegan and Clifford, 2001). Additionally, the use of systems without water exchange is a measure that reduces the risk of disease introduction (Pruder, 2004; Hasan et al., 2020).
However, Fegan and Clifford (2001) and OIE (2016) state that although water may act as an important vehicle for the spread of viruses in the aquatic environment, a high viral load is required for the animal to become infected and ill. Water transmission of WSSV has been demonstrated in different experiments. However, most of them were done with high viral titrations or unnatural proximity of infected to uninfected animals. In our study, crabs were collected inside and in the vicinity of the shrimp farms and their viral loads were measured by qPCR. Although data in the literature demonstrate high mortality as a consequence of WSSV infection, it appears to be dependent on both free virus concentration in water and the health status of exposed animals. Along with the fact that the virus does not remain viable outside a host for more than a few days, it indicates that the risk of water transmitting WSSV is considered lower than previously believed, except when water is discharged with a high viral load during a period of outbreak (Fegan and Clifford, 2001). It is not clear whether crabs were infected by WSSV-positive farmed shrimp or, on the contrary, were responsible to spread the virus to shrimps. However, once crabs are asymptomatic vector of WSSV, an understanding of the reasons for differences in mortality from equally heavy viral loads may allow us to develop strategies for limiting mortality from viral pathogens in shrimp aquaculture (Kanchanaphum et al., 1998; Marques et al., 2011; Bateman and Stentiford, 2017).
So far, evidences regarding whether N. granulata crabs show some kind of resistance to WSSV due to the ability of either clear out the virus, inactivate it, or even slow down its multiplication is not yet possible to say. We observed that equal viral loads promote differences in some molecular responses between shrimp and crabs, as seen in the transcript level of some target genes. So apparently crabs may modulate some molecular pathways or adaptive strategies to avoid the drastic effects of WSSV infection, while shrimp succumb faster to this viral pathogen. Comparatively, viral loads, in both species, were different along the course of infection. WSSV replicated more rapidly in shrimp when compared with crabs. Moreover, our results show that in general terms crabs were less susceptible to WSSV.
4.2. Differential expression in N. granulata and L. vannamei related to the same viral load
Our gene transcript findings showed statistical differences in crabs and shrimp at the same viral load, pointing out similar potential effects of WSSV infection in molecular processes in both species, such as apoptosis, ions storage, and involvement of chaperone proteins. Along the progress of infection, a wide range of molecular interactions occur between WSSV and its host cells. These molecular interactions play an important role in determining host susceptibility to the pathogen.
Verbruggen et al. (2016) have listed a variety of molecular mechanisms that respond and even prevent WS host infectivity. These mechanisms involve several pathways, including some of genes we targeted in the present study. Genes associated with chaperone and apoptosis pathways, such as CALC, IAP, HSP 70, H2a and UBQ, and others associated with phenoloxidase activation, such as PHEN, QM and FERR were cleared induced in both, shrimp and crabs. Upon reaching the threshold of 106 viral copies, all the shrimp died while the crabs remained alive, showing no visible signs of disease. Despite the antiviral responses caused by the shrimp cells, this molecular defense was not enough to keep the host alive.
Apoptosis is considered an important cellular defense mechanism that inhibits viral multiplication and eliminates infected cells in multicellular organisms (Everett and McFadden, 1999). Leu et al. (2013) propose a model for apoptotic interaction between WSSV and shrimp through the activation of signaling pathways that lead to (1) the expression of pro-apoptosis proteins, like caspase modulators; and (2) mitochondrial changes. In a few cases where viruses intentionally induce apoptosis to release progeny virus, the inhibitor of apoptosis proteins (IAP) plays an important role in both apoptosis and innate immunity. The IAP proteins are considered as strict regulators of caspase/apoptosis activity and are influenced by viral infections. Kulkarni et al. (2014) state that in Penaeus monodon when IAP expression was present, caspases level decrease. Comparative analyses in our study, between WSSV-challenged groups, showed a decrease of CASP-3 levels in WSSV-infected shrimp, and a decrease of CASP-2 and CASP-3 levels in WSSV-infected crabs. To manipulate host apoptosis, WSSV modulates the expression of apoptosis-related genes, such as caspase and fortilin, as described to shrimp Marsupenaeus japonicus, Penaeus monodon and to crab Eriocheir sinensis and Cherax quadricarinatus, to actively promote apoptosis to spread virus progeny to neighboring cells (Wang et al., 2013; Qu et al., 2018; Li et al., 2019).
Further, HSP60 is considered a pro-apoptotic protein, whereas HSP27, HSP70, and HSP90 proteins are predominantly anti-apoptotic (Murthy and Ravishankar, 2016). In L. vannamei, HSP family was consistently or specifically expressed in response to thermal or pH stress, heavy metal exposure (Qian et al., 2012), besides viral infection stress. According to Valentim-Neto et al. (2014a), shrimp positive to Hypodermal and Hematopoietic Necrosis Virus (IHHNV) or WSSV with changes in HSP 70 expression levels had a higher rate of survival during the period of cultivation. On the other hand, we observed that in crab and shrimp with severe WSSV infection, transcripts of HSP 70 showed to be up-regulated, as expected, since a role in shrimp antiviral response has been attributed to this gene (Moser and Valentim-Neto, 2020). Perhaps viral infections, such as that caused by WSSV, may cause similar tissue and protein damages as those attributed to heat shock stress. On the other hand, the up-regulation of HSP 70 genes may act to repair protein damages and may play an additional role as signaling molecules to modulate the innate immune response in host shrimp (Janewanthanakul et al., 2019). We also observed a decrease in HSP60 transcript at 24 to 48 hpi, when the WSSV load raised from 101 to 106 copies.µl− 1. In crabs, WSSV-infection increased slower between 96 to 120 hpi. Sun et al. (2013) also report that HSP 60 expression inhibited in gills of WSSV-infected L. vannamei. Zhou et al. (2010) reported a significant HSP 60 expression in L.vannamei gills after Vibrio alginolyticus challenge followed by a decrease to normal levels after 24 h.
The transcript levels of ubiquitin (UBQ) were up-regulated in WSSV infected shrimp and crabs at the same viral load. Our results also showed that levels of ubiquitin transcripts varied depending on the WSSV post-infection time interval, as well as the viral load, both in cabs and shrimp. Vidya et al. (2013) suggested that one of the viral strategies to keep progressive infection involves modifying host ubiquitination. Protein degradation pathway is the most well-studied aspect of the ubiquitin-proteasome system; protein ubiquitination is also responsible for regulating cell signaling by controlling the endocytosis. In addition to the protein degradation pathway, ubiquitin also participates in other cell functions, such as activation of immune cells (Ben-Neriah, 2002) and apoptosis (Leu et al., 2013). As ubiquitin is a protein that acts in several processes the data found in the current literature refer to both a viral pro-infection function induced by WSSV, as well as a possible anti-viral function produced by the host itself. An induction of UBQ has also been seen in other studies of shrimp infected with WSSV (He et al., 2009; Wang et al., 2006 a). Levels of ubiquitin were up-regulated in WSSV-challenged L.vannamei after 24 hpi in comparison to non-infected shrimp, whereas after 72 hpi the same protein was present only in infected shrimp (Valentim-Neto et al., 2014a). Viral load was also monitored in the same study and ranged from 8.38x101 (24 hpi) to 7,79x105 copies (72 hpi). Further, Chen et al. (2016) report that in shrimp P. monodon injected with two viral proteins with the purpose of gaining resistance against WSSV, the list of up-regulated protein spots found exclusively in WSSV-vaccinated shrimp included ubiquitin, calreticulin and HSP 70. All together, these findings show the important role of ubiquitin along the WSSV infection, as well as the involvement of stress proteins and proteins related to calcium metabolism, calreticulin (Chen et al., 2016) and calcitonin-like (Valentim-Neto et al., 2014a).
The histone H2ɑ has been recently implicated in the apoptosis process and identified among the participant proteins in WSSV infection studies (Encinas et al., 2019; Wang et al., 2008). The histone H2ɑ is responsible for the packaging and compactation of nuclear DNA and plays an important role during the regulation of genes at the nuclear level. In addition, H2ɑ displays a high diversity of variants that are involved in apoptosis DNA repair, gene regulation, and genome integrity (González-Romero et al., 2012). Wang et al. (2008) showed that ICP11, a WSSV protein, bound with histone proteins in the cytosol of WSSV-infected hemocytes and HeLa cells, prevent them to participate in nucleosome assembly, inducing incidental apoptosis. Our results show that H2ɑ gene is up-regulated in crabs and shrimp at the same viral load. But, interestingly enough, we observed a significant higher transcription before viral load achieve such level (106 copies.µl− 1), that is, at 24 hpi in shrimp and at 96 hpi in crabs. These results corroborated with Encinas et al. (2019) that reported that maximum levels of expression of this protein were reached at 12h post-WSSV-infection in L. vannamei, declining sharply after that. Feng et al. (2014a) observed that H2ɑ transcript levels in WSSV-challenged Fenneropenaeus chinensis displayed peaks of expression at 6 and 36 hpi. These results indicate an early response of histone protein against WSS|V, as long as the viral infection is considered mild or moderate.
Under severe WSSV-infection our study identified the up-regulation of QM in crabs, as in shrimp further implicating the role of the product of this gene in defense reactions in crustaceans. Xu et al. (2008) demonstrated that the M. japonicus QM protein could regulate the phenoloxidase activity by interaction with hemocyanin, suggesting the involvement of QM protein in the prophenoloxidase (proPO) activation system in shrimp immunity. Liu et al. (2014) report that QM transcripts in L. vannamei were significantly increased after challenge with Vibrio anguillarum. Likewise, an up-regulated expression on phenoloxidase (PHEN) was observed in our study in response to WSSV in both crustacean species. On the other hand, the expression of proPO in the WSSV-infected shrimp was not significant, even considering that proPO was primarily expressed in hemocytes and many tissues infiltrated by hemocytes, like gills, and has been well described as shrimp viral defense according to several reports (Burnett and Burnett, 2015; Wang et al., 2006b; van de Braak et al., 2002). On the contrary, our study detected statistical differences in proPO gene in WSSV-infected crabs. Thus, apparently high viral load induced QM and PHEN gene transcript levels in both challenged crustacean species but was not able to active the whole pro-phenoloxidase system.
Different research results have confirmed the existence of a correlation between iron metabolism and the immune system in crustaceans, once iron is a co-factor of ribonucleotide reductase and is necessary for enzyme activity (Zhang et al., 2014). The depletion of cellular iron can lead to the inhibition of ribonucleotide reductase, preventing virus proliferation (Lin et al., 2015; Verbruggen et al., 2016). Ferritin is a major iron storage protein in living cells and plays an important role in iron homeostasis and also in host's innate immune response to various pathogens (Ye et al., 2015). Our findings revealed up-regulated ferritin gene expression (FERR) in crabs and shrimp, in response to severe WSSV infection. In invertebrates, FERR was found to be up-regulated after pathogens challenge and is considered to be an important element in the innate immune system. The transcripts of FERR in shrimp gills were reported to be up-regulated post WSSV challenge by Ye et al. (2015) once the expression of ferritin was significantly increased 12 h after WSSV injection and was kept in high level untill 96 h after WSSV injection in shrimp. In addition, the up-regulation of ferritin has also been observed in WSSV-infected M. japonicus (Feng et al., 2014b) and in C. quadricarinatus (Chen et al., 2018).
The transcripts of calreticulin (CALR), a highly conserved endoplasmic reticulum luminal resident protein, were up-regulated in shrimp and down-regulated in crab gills at the same viral charge. In crustaceans, calreticulin is known to play important roles in in calcium homeostasis, molting, immune functions, and stress response to viral infection (Luana et al., 2007; Huang et al., 2019). Also, the calcified cuticle proteins (CALC) are related to the cuticle synthesis in crustaceans, with their transcription increased during the molting phase (Kuballa et al., 2007). This protein has never been directly related to WSSV infection, prior to Müller studies (2009), but its induction may be related to the abnormal deposit of calcium salts in the cuticle of the animals, which characterizes WSD. In our experiment, CALC transcript levels were up-regulated in severe WSSV infection, both in crabs and shrimp. It’s well established that shrimp acutely infected with WSSV often show abnormal deposits of calcium within the cuticle (Wang et al., 2006), as well as soft cuticles in advances stages of infection, signs that may be related with the modulation of transcripts encoding to CALR and CALC genes.
Decapod crustaceans (marine and freshwater) are susceptible hosts to the development of the WSD, while non-decapod crustaceans can accumulate high concentrations of viral particles without proof of viral replication in these organisms. However, this is the first report of experimental infection by WSSV and viral proliferation in N. granulata crabs. The set of genes investigated in our study could be used as a complementary early-warning biomarkers to monitor the health status and susceptibility of shrimp and other crustaceans to WSSV, since these responses are directly related to the viral infection. Additionally, the same genes could also be analyzed in early phases or along the time course of the infection to better understand the dynamics of WSSV infection, in shrimp and crabs, and better study the difference in susceptibility to the infection.
A better knowledge of the molecular defense related to host antiviral responses may contribute to understand and explore the mechanisms triggered by WSSV. The absence of mortality or clinical signs, as seen in N. granulata, may provide the virus a better opportunity for replication and efficient continuing transmission. Moreover, the passage of a WSSV strain through different hosts induces genomic variation and alters the pathogenicity of the virus (Waikhom et al., 2006; Müller et al., 2010), which could explain the variation in WSSV-stress responses. A better understanding of the reasons, as well the molecular scenario, for differences in mortality at equally heavy viral infections may allow us to develop strategies for limiting mortality from viral pathogens in shrimp aquaculture.
In conclusion, our investigation provides evidence that N. granulata is susceptible to WSSV and the virus is capable to replicate into its cells. WSSV-challenge was conducted by intramuscular injection and revealed that N. granulata is less susceptive to WSSV than L. vannamei. The WSSV-stress caused by viral load increase affected transcript levels of eleven genes in crabs and six genes in shrimp, modulating different cellular mechanisms, such as apoptosis, iron storage and mobilization, and chaperone stresse response. These molecular interaction network play an important role in determining host susceptibility and infectability of the pathogen.
Funding information
This work was supported by the Brazilian official funding agency, The National Council for Scientific and Technological Development – CNPq, Proc.485054/2012-4 and Proc. 311099/2013-0
- Balasubramanian, C.P., Anand, S., Kannappan, S., Biju, I.F., 2018. Training manual on recent advances in farming of pacific white shrimp (Penaeus vannamei). Training manual series, n 14, 126 pp.
- Ben-Neriah, Y., 2002. Regulatory functions of ubiquitination in the immune system. Nat. Immunol. 3, 20-26. https://doi.org/10.1038/ni0102-20
- Blaylock, R.B., Curran, S.S., Lotz, J.M., 2019. White spot syndrome virus (WSSV) in cultured juvenile blue crabs Callinectes sapidus: oral versus injection exposure, and feeding frequency effects. Dis. Aquat. Org. 133, 147-156. https://doi.org/10.3354/dao03334
- Bateman, K.S., Stentiford, G.D., 2017. A taxonomic review of viruses infecting crustaceans with an emphasis on wild hosts. Journal of Invertebrate Pathology, 147, 86–110. https://doi:10.1016/j.jip.2017.01.010
- van de Braak, C.B., Botterblom, M.H., Huisman, E.A., Rombout, J,H., van der Knaap, W.P., 2002. Preliminary study on haemocyte response to white spot syndrome virus infection in black tiger shrimp Penaeus monodon. Diseases of Aquatic Organisms, 51(2):149-155. https://doi: 10.3354/dao051149.
- Burnett, K. G., Burnett, L. E. 2015. Respiratory and Metabolic Impacts of Crustacean Immunity: Are there Implications for the Insects? Integrative and Comparative Biology, 55(5), 856–868. https://doi: 10.1093/icb/icv094
- Cavalli, L.S, Batista, C.R., Nornberg, B.F., Mayer, F.Q., Seixas, F.K., Romano, L.A., Marins, L.F., Abreu, P.C., 2013. Natural occurrence of White spot syndrome virus and Infectious hypodermal and hematopoietic necrosis virus in Neohelice granulata crab. J. Invertebr Pathol. 114, 86-88. https://doi.org/10.1016/j.jip.2013.06.002.
- Chen, L.H., Lin, S.W., Liu, K.F., Chang, C.I., Hseu, J.R., Tsai, J.M., 2016. Comparative proteomic analysis of Litopenaeus vannamei gills after vaccination with two WSSV structural proteins. Fish Shellfish Immunol. 49, 306-314. https://doi.org/10.1016/j.fsi.2015.12.044.
- Chen, X.X., Li, Y.Y., Chang, X.J., Xie, X.L., Liang, Y.T., Wang, K.J., Liu, H.P., 2018. A CqFerritin protein inhibits white spot syndrome virus infection via regulating iron ions in red claw crayfish Cherax quadricarinatus. Dev. Comp. Immunol. 82, 104-112. https://doi.org/10.1016/j.dci.2018.01.008.
- de Souza Valente, C.; Ortiz, K.O.; Fraga, A.P.M.; Vieira, F. N.; Marques, M.R.F., 2020. Transcription of defense related genes in Pacific white shrimp, Litopenaeus vannamei, kept in biofloc and clear seawater and challenged with the white spot syndrome virus. Aquaculture International, 28, 293-307. https://doi.org/10.1007/s10499-019-00461-8.
- Durand, S., Lightner, D.V., Redman, R.M, Bonami, J.R., 1997. Ultrastructure and morphogenesis of white spot syndrome baculovirus (WSSV). Dis. Aquat. Org. 29, 205-211. https://doi.org/10.1006/viro.2001.1002.
- Encinas-García, T., Loreto-Quiroz, D.L., Mendoza-Cano, F., Peña-Rodriguez, A., Fimbres-Olivarria, D., de la Vega, E., Sánchez-Paz, A., 2019. White spot syndrome virus down-regulates expression of histones H2A and H4 of Penaeus vannamei to promote viral replication. Dis. Aquat. Org. 137, 73-79. https://doi.org/10.3354/dao03428.
- Everett, H., McFadden, G., 1999. Apoptosis: an innate immune response to virus infection. Trends Microbiol. 7(4), 160–165. https://doi.org/10.1016/s0966-842x(99)01487-0.
- Fegan, D.; Clifford III, H.C., 2001. Health management for viral diseases in shrimp farms, in: Browdy, C.L.; Jory, D.E. (Eds.), The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture, The World Aquaculture Society, Baton Rouge, LA, pp. 168-198.
- Feng, J., Tang, X., Zhan, W., 2014a. Analysis and identification of tyrosine phosphorylated proteins in hemocytes from Fenneropenaeus chinensis (Decapoda: Penaeidae) infected with white spot syndrome virus. J. Crustacean Biol. 34, 453-459. https://doi.org/10.1163/1937240x-00002241
- Feng, W.R., Zhang, M., Su, Y.Q., Wang, J., Wang, Y.T., Mao, Y., 2014b. Identification and analysis of Marsupenaeus japonicus ferritin that is regulated at the transcriptional level by WSSV infection. Gene. 544:184e90. https://doi.org/10.1016/j.gene.2014.04.05.
- Flegel, T.W., 2006. Detection of major penaeid shrimp viruses in Asia, a historical perspective with emphasis on Thailand. Aquaculture. 258, 1–33. https://doi.org/10.1016/j.aquaculture.2006.05.013
- González-Romero, R., Rivera-Casas, C., Frehlick, L.J., Méndez, J., Ausió, J., Eirín-López, J.M., 2012. Histone H2A (H2A.X and H2A.Z) Variants in Molluscs: Molecular characterization and potential implications for chromatin dynamics. PLoS ONE, 7:e30006. https://doi.org/10.1371/journal.pone.0030006
- Goularti Filho, A., Ronçani, L. D., 2018. Carcinicultura em Santa Catarina: da euforia desregulada à crise generalizada. Historia Ambiental Latinoamericana Y Caribeña (HALAC) Revista De La Solcha. 8, 67-91. https://doi.org/10.32991/2237-2717.2018v8i1.p67-91
- Hasan, N. A., Haque, M. M., Hinchliffe, S. J., Guilder, J., 2020. A sequential assessment of WSD risk factors of shrimp farming in Bangladesh: Looking for a sustainable farming system. Aquaculture. 26, 735348. https://doi:10.1016/j.aquaculture.2020.735348
- He, F., Syed, S.M., Hameed, A., Kwang, J., 2009. Viral ubiquitin ligase WSSV222 is required for efficient white spot syndrome virus replication in shrimp. J. Gen Virol. 90, 1483-1490. https://doi.org/10.1099/vir.0.008912-0.
- Huang, H., Huang, C., Guo, L., Zeng, C., Ye, H., 2019. Profiles of calreticulin and Ca2+ concentration under low temperature and salinity stress in the mud crab, Scylla paramamosain. PLoS ONE 14(7): e0220405. https://doi.org/10.1371/journal.pone.0220405.
- Janewanthanakul, S., Supungul, P., Tang, S., Tassanakajon, A., 2019. Heat shock protein 70 from Litopenaeus vannamei (LvHSP70) is involved in the innate immune response against white spot syndrome virus (WSSV) infection. Developmental & Comparative Immunology. 103476. https://doi:10.1016/j.dci.2019.103476
- Kanchanaphum, P., Wongteerasupaya, C., Sitidilokratana, N., Boonsaeng, V., Panyim, S., Tassanakajon, A., Withyachumnarnkul, B., Flegel, T.W., 1998. Experimental transmission of white spot syndrome virus (WSSV) from crabs to shrimp Penaeus monodon. Dis. Aquat. Org. 34, 1-7. https://doi.org/10.3354/dao03400.
- Kuballa, P., Matentzoglu, K., Scheffner, M., 2007. The role of the ubiquitin ligase E6-AP in human papillomavirus E6-mediated degradation of PDZ domain-containing proteins. J. Biol. Chem. 282, 65-71. https://doi.org/10.1074/jbc.m605117200.
- Kulkarni, A.D., Caipang, C.M.A., Kiron, V., Rombout, J.H.W.M., Fernandes, J.M.O., Brinchmann, M.F., 2014. Evaluation of immune and apoptosis related gene responses using an RNAi approach in vaccinated Penaeus monodon during oral WSSV infection. Mar. Genom. 18, 55–65. https://doi.org/10.1016/j.margen.2014.05.002.
- Leu, J.H., Lin, S.J., Huang, J.Y., Chen, T.C., Lo, C.F., 2013. A model for apoptotic interaction between white spot syndrome virus and shrimp. Fish Shellfish Immunol. 34, 1011-1017. https://doi.org/10.1016/j.fsi.2012.05.030.
- Li, Y.Y., Xie, X.L., Ma, X.Y., Liu, H.P., 2019. Identification of a CqCaspase gene with antiviral activity from red claw crayfish Cherax quadricarinatus. Dev. Comp. Immunol. 91, 101-107. https://doi.org/10.1016/j.dci.2018.10.012.
- Lin, S.J., Lee, D.Y., Wang, H.C., Kang, S.T., Hwang, P.P., Kou, G.H., Huang, M.F., Chang, G.D., Lo, 471 C.F., 2015. White spot syndrome virus protein kinase 1 defeats the host cell's iron-withholding defense 472 mechanism by interacting with host ferritin. J. Virol. 89, 1083-1093. https://doi.org/10.1128/JVI.02318-14.
- Liu, W., Qian, D., Yan, X. J., 2011. Studies on pathogenicity and prevalence of white spot syndrome virus in mud crab, Scylla serrata (Forskal), in Zhejiang Province, China. J. Fish Dis. 34, 131-138. https://doi.org/10.1111/j.1365-2761.2010.01221.x.
- Liu, Y., Qian, Z., Qu, R., Wang, X., He, S., Hou, F., Liu, Q., Mi, X., Liu, X., 2014. Molecular characterization, immune responsive expression and functional analysis of QM, a putative tumor suppressor gene from the Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 37, 1-10. https://doi.org/10.1016/j.fsi.2014.01.005.
- Lo, C.F., Ho, C.H., Chen, C.H., Liu, K.F., Chiu, Y.L., Yeh, P.Y., Peng, S.E., Hsu, H.C., Liu, H.C., Chang, C.F., Su, M.S., Wang, C.H., Kou, G.H., 1997a. Detection and tissue tropism of White spot syndrome baculovirus (WSBV) in captured brooders of Penaeus monodon with a special emphasis on reproductive organs. Dis. Aquat. Org. 30, 53–72. https://doi.org/10.3354/dao030053.
- Lo, C.F., Ho, C.H., Peng, S.E., Chen, C.H., Hsu, H.C., Chiu, Y.L., Chang, C.F., Liu, K.F., Su, M.S., Wang, C.H. and Kou, G.H. 1996a: White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Org. 27, 215–225.
- Lo, C.F., Leu, J.H., Ho, C.H., Chen, C.H., Peng, S.E., Chen, Y.T., Chou, C.M., Yeh, P.Y., Huang, C.J., Chou, H.Y., Wang, C.H., Kou, G.H., 1996b. Detection of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimps using polymerase chain reaction. Dis Aquat Organ. 25, 133-141. https://doi.org/10.3354/dao025133.
- Luana, W., Li, F., Wang, B., Zhang, X., Liu, Y., Xiang, J., 2007. Molecular characteristics and expression analysis of calreticulin in Chinese shrimp Fenneropenaeus chinensis. Comp Biochem Physiol B Biochem Mol Biol. 147, 482–491. https://doi.org/10.1016/j.cbpb.2007.03.001.
- Maeda, M., Itami, T., Furumoto, A., Hennig, O., Imamura, T., Kondo, M., Hirono, I., Aoki, T., Takahashi, Y., 1998. Detection of penaeid rod-shaped DNA virus (PRDV) in wild-caught shrimp and other crustaceans. Fish Pathol. 33, 373-380. https://doi.org/10.3147/jsfp.33.373.
- Marques, J.S., Müller, I.C., Moser, J.R., Sincero, T.C., Marques, M.R.F., 2011. Wild captured crab, Chasmagnathus granulata (Dana, 1851), a new host for white spot syndrome virus (WSSV). Aquaculture. 318, 20 – 24. https://doi.org/10.1016/j.aquaculture.2011.04.031.
- Mayo, M.A., 2002. A summary of taxonomic changes recently approved by ICTV. Arch Virol. 14, 1655–1656. https://doi.org/10.1007/s007050200039.
- Melo, G.A.S., 1996. Manual de identificação dos Brachyura (caranguejos e siris) do litoral brasileiro. Plêiade/FAPESP Ed., São Paulo, 604p.
- Meng, X.H.; Jang, I.K., Seo, H. C, Cho, Y.R., 2009. White spot syndrome virus quantification in blue crab Portunus trituberculatus hatchery-produced larvae and wild populations by TaqMan real-time PCR, with an emphasis on the relationship between viral infection and crab health. Aquaculture. 291,18–22. https://doi.org/10.1016/j.aquaculture.2009.02.003.
- Mijangos-Alquisires, Z., Quintero-Arredondo, N., Castro-Longoria, R., Grijalva-Chon, J. M., Ramos-Paredes, J., 2006. White spot syndrome virus (WSSV) in Litopenaeus vannamei captured from the Gulf of California near an area of extensive aquaculture activity. Dis. Aquat. Org. 71, 87–90. https://doi.org/10.3354/dao071087.
- Moser, J.R.; Álvarez, D. A. G.; Cano, F. M.; Encinas Garcia, T.; Molina, D.E. C.; Clark, G. P.; Marques, M.R. F.; Magallón, F.J.B.; Hernández López, J., 2012. Water temperature influences viral load and detection of White Spot Syndrome Virus (WSSV) in Litopenaeus vannamei and wild crustaceans. Aquaculture. 326-329, 9-14.
- Moser, J. R., Valentim-Neto, P. A., 2020. Heat Shock Proteins HSP 70: a review on shrimps. In: Rajib Deb; N.V. Patil; Gyanendra Singh Sengar. (Org.). HSP70s: discovery, structure and functions. 1ed.New York: Nova Science Publishers. 37-92.
- Müller, I.C., 2009. Genes diferencialmente expressos em camarões de cultivo Litopenaeus vannamei infectados pelo vírus da Síndrome da Mancha Branca e genotipagem de isolados geográficos brasileiros do vírus. Tese de Doutorado. Programa de Pós-Graduação em Biotecnologia e Biociências. Universidade Federal de Santa Catarina – UFSC.
- Müller, I.C., Andrade T.P.D., Tang-Nelson K.F.J., Marques M.R..F, Lightner D.V. 2010.
Genotyping of white spot syndrome virus (WSSV) geographical isolates from Brazil and comparison to other isolates from the Americas. Dis. Aquat. Org. 88, 91-98. https://doi.org/10.3354/dao02142. - Murthy, V.S., Ravishankar, K.V., 2016. Molecular mechanisms of Heat Shock Proteins and thermotolerance in plants, in: Srinivasa Rao, N.K., Shivashankara, K.S., Laxman, R.H. (Eds.), Abiotic stress physiology of horticultural crops. Springer, India, Nova Deli. pp. 71–83. https://doi.org/10.1007/978-81-322-2725-0_4.
- Nunan, L. M., Lightner, D. V., 2011. Optimized PCR assay for detection of white spot syndrome virus (WSSV). Journal of Virological Methods. 171(1), 318–321. https://doi:10.1016/j.jviromet.2010.11.015
- Oidtmann, B., Stentifordm G.D., 2011. White Spot Syndrome Virus (WSSV) concentrations in crustacean tissues – A review of data relevant to assess the risk associated with commodity trade. Transbound Emerg. Dis. 58, 469–482. https://doi.org/10.1111/j.1865-1682.2011.01231.x.
- OIE – World Organization for Animal Health. 2016. Manual of Diagnostic Tests for Aquatic Animals, Office International des Epizooties, Paris, France. 7th Edition, 589 pp.
- Peng, S.E., Lo C.F., Ho, C.H., Chang, C.F., Kou, G.H., 1998. Detection of white spot baculovirus (WSBV) in giant freshwater prawn, Macrobrachium rosenbergii, using polymerase chain reaction. Aquaculture. 164, 253-262. https://doi.org/10.1016/S0044-8486(98)00191-4.
- Pruder, G.D., 2004. Biosecurity: application in aquaculture. Aquacult. Eng. 32, 3-10. doi:10.1016/j.aquaeng.2004.05.002.
- Qian, Z., Liu, X., Wang, L., Wang, X., Li, Y., Xiang, J., Wang, P., 2012. Gene expression profiles of four heat shock proteins in response to different acute stresses in shrimp, Litopenaeus vannamei. Comp Biochem Physiol C Toxicol Pharmacol. 156, 211-220. doi:10.1016/j.cbpc.2012.06.001.
- Qu, C., Yang, W., Xu, Q., Sun, J., Lu, M., Wang, Y., Liu, C., Wang, W., Wang, L., Song, L., 2018. A novel effector caspase (Caspase-3/7-1) involved in the regulation of immune homeostasis in Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 83, 76-83. https://doi.org/10.1016/j.fsi.2018.09.013.
- Rahman, M. Meezanur, Mathias Corteel, Mathieu Wille, Victoria Alday-Sanz, Maurice B. Pensaert, Patrick Sorgeloos, and Hans J. Nauwynck., 2007. The effect of raising water temperature to 33 °C in Penaeus vannamei juveniles at different stages of infection with White Spot Syndrome Virus (WSSV). Aquaculture. 272, 240–245. https://doi.org/10.1016/j.aquaculture.2007.07.228.
- Sánchez-Paz, A., Terán-Díaz, B., Enríquez-Espinoza, T., Encinas-Garcia, T., Vázquez-Sánchez, I., & Mendoza-Cano, F., 2015. The tidepool shrimp, Palaemon ritteri Holmes, constitutes a novel host to the white spot syndrome virus. J. Fish Dis. 38, 613–620. https://doi.org/10.1111/jfd.12275.
- Sahul Hameed, A.S., Balasubramanian, G., Musthaq, S.S., Yoganandhanl, K., 2003. Experimental infection of twenty species of Indian marine crabs with white spot syndrome virus (WSSV). Dis. Aquat. Org. 57, 157-161. https://doi.org/10.3354/dao057157.
- Sahul Hameed, A.S., Yoganandhanl, K., Sathish, S., Rasheed, M., Murugan, V., Jayaraman, K., 2001. White spot syndrome virus WSSV in two species of freshwater crabs Paratelphusa hydrodomous and P. pulvinata. Aquaculture. 201, 179-186. https://doi.org/10.1016/S0044-8486(01)00525-7.
- Sakai, K., Türkay, M., Yang, S.L., 2006. Revision of the Helice/Chasmagnathus complex (Crustacea: Decapoda: Brachyura). Abh Senckenberg Naturforsch Ges. 565, 1–76.
- Schmittgen, T. D., Livak, K. J., 2008. Analyzing real-time PCR data by the comparative CT method. Nature Protocols. 3(6), 1101–1108. https://doi:10.1038/nprot.2008.73
- Shekhar, M.S., AZAD, I.S., Ravichandran, P., 2006. Comparison of dot blot and PCR diagnostic for detection of white spot syndrome virus in different tissues of Penaeus monodon. Aquaculture. 261, 1122-1127.
- Seiffert, W., Costa, S.W., Maggioni, D.A., 2005. Mancha branca em Santa Catarina. Revista Panorama da Aquicultura. 15, 51-53.
- Souza, T.M., 2008. Detecção do vírus da mancha branca (WSV) em Litopenaeus vannamei por métodos de diagnóstico moleculares: hibridização in situ e PCR em tempo real. Dissertação (mestrado) - Universidade Federal de Santa Catarina, Centro de Ciências Agrárias. Programa de Pós-Graduação em Aquicultura.
- Spivak, E.D., 2010. The crab Neohelice (=Chasmagnathus) granulata: An emergent animal model from emergent countries. Helgol Mar Res. 64, 149-154. https://doi.org/10.1007/s10152-010-0198-z.
- Sun, F., Liu, Q.H., Huang, J., 2013. Expression of HSP60 and HSP90 in shrimp Litopenaeus vannamei during WSSV infection. (Chinese J) Progress in Fishery Sciences, 34 (2): 90 - 95
- Suttler, C.A., 2005. Viruses in the sea. Nature. 437, 356 - 361. https://doi.org/10.1038/nature04160.
- Valentim-Neto, P.A., Fraga, A. P., Marques, M.R.F., 2014a. Differential expression of proteins in the gills of Litopenaeus vannamei infected with white spot syndrome virus. Aquaculture International. 22, 1605-1620. https://doi.org/10.1007/s10499-014-9768-4.
- Valentim-Neto, P.A., Fraga, A.P.M., Müller, G.A.S., Marques, M.R.F., 2015. Protein expression profiling in the gill of Litopenaeus vannamei (Boone, 1931) naturally infected with white spot syndrome virus. Crustaceana. 88, 747-765. https://doi.org/10.1163/15685403-00003446.
- Valentim-Neto, P.A., Moser, J.R., Fraga, A.P.M., Marques, M.R.F., 2014b. HSP70 expression in shrimp Litopenaeus vannamei in Response to IHHNV and WSSV Infection. Virus Dis. 25, 437-440. https://doi.org/10.1007/s13337-014-0236-6.
- van Hulten, M.C.W., Witteveldt, J., Peters, S., Kloosterboer, N., Tarchini, R., Fiers, M., Sandbrink, H., Lankhorst, R.K., Vlak, J.M., 2001. The white spot syndrome virus DNA genome sequence. Virology. 286, 7–22. https://doi.org/10.1006/viro.2001.1002.
- Verbruggen, B., Bickley, L.K., Van Aerle, R., Bateman, K.S., Stentiford, G.D., Santos, E.M., Tyler, C.R., 2016. Molecular mechanisms of White Spot Syndrome Virus infection and perspectives on treatments. Viruses. 8, 23. https://doi.org/10.3390/v8010023.
- Vidya, R., Gireesh-Babu, P., Pani Prasad, K., 2013. White Spot Syndrome Virus manipulates ubiquitin gene expression in Penaeus monodon. Indian J. Virol. 24, 82–84. https://doi.org/10.1007/s13337-012-0113-0.
- Vijayan, K.K., Stalin Raj, V., Balasubramanian, C.P., Alavandi, S.V., Thillai Sekhar, V., Santiago, T.C., 2005. Polychaete worms - a vector for white spot syndrome virus (WSSV). Dis. Aquat. Org. 63, 107-111. https://doi.org/10.3354/dao063107.
- Waikhom, G., John, K.R., George, M.R., Jeyaseelan, M.J.P., 2006. Differential host passaging alters pathogenicity and induces genomic variation in white spot syndrome virus. Aquaculture. 261, 54–63. https://doi.org/10.1016/j.aquaculture.2006.07.031.
- Walker, P. J., Gudkovs, N., Mohan, C. V., Raj, V. S., Pradeep, B., Sergeant, E., Chandra Mohan, A.B., Ravibabu, G., Indrani Karunasagar, Santiago, T. C. 2011. Longitudinal disease studies in small-holder black tiger shrimp (Penaeus monodon) ponds in Andhra Pradesh, India. II. Multiple WSSV genotypes associated with disease outbreaks in ponds seeded with uninfected postlarvae. Aquaculture, 319(1-2), 18–24. https://doi:10.1016/j.aquaculture.2011.06.021
- Wang, B., Li, F., Dong, B., Zhang, X., Zhang, C., Xiang, J., 2006 a . Discovery of the genes in response to white spot syndrome virus (WSSV) infection in Fenneropenaeus chinensis through cDNA microarray. Mar. Biotechnol. 8, 491-500. https://doi.org/10.1007/s10126-005-6136-4.
- Wang, Y.C., Chang, P.S., Chen, H.Y. 2006 b. Tissue distribution of prophenoloxidase transcript in the Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 20(3):414-8. https://doi: 10.1016/j.fsi.2005.05.003. PMID: 16039880.
- Wang, H.C., Wang, H.C., Ko, T.P., Lee, Y.M., Leu, J.H., Ho, C.H., Wang, A.H.J., 2008. White spot syndrome virus protein ICP11: A histone-binding DNA mimic that disrupts nucleosome assembly. PNAS. 105, 20758 – 20763. https://doi.org/10.1073/pnas.0811233106.
- Wang, P.H., Wan, D.H., Gu, Z.H., Qiu, W., Chen, Y.G., Weng, S.P., Yu, X.Q., He, J.G., 2013. Analysis of expression, cellular localization, and function of three inhibitors of apoptosis (IAPs) from Litopenaeus vannamei during WSSV Infection and in regulation of antimicrobial peptide genes (AMPs). PLoS ONE 8(8): e72592. https://doi.org/10.1371/journal.pone.0072592.
- Wang, Y.C., Lo, C.F., Chang, P.S., Kou, G.H., 1998. Experimental infection of white spot baculovirus in some cultured and wild decapods in Taiwan. Aquaculture. 164, 221-231. https://doi.org/10.1016/S0044-8486(98)00188-4
- Xu, J., Wu, S., Zhang, X., 2008. Novel function of QM protein of shrimp (Penaeus japonicus) in regulation of phenol oxidase activity by interaction with hemocyanin. Cell Physiol Biochem. 21, 473–480. https://doi.org/10.1159/000129640.
- Yan, D.C., Dong, S.L., Huang, J., Yu, X.M., Feng, M.Y., Liu, X.Y., 2004. White spot syndrome virus (WSSV) detected by PCR in rotifers and rotifer resting eggs from shrimp pond sediments. Dis. Aquat. Org. 59, 69–73. https://doi.org/10.3354/dao059069.
- Ye, T., Wu, X., Wu, W., Dai, C., Yuan, J., 2015. Ferritin protect shrimp Litopenaeus vannamei from WSSV infection by inhibiting virus replication. Fish Shellfish Immunol. 42, 138–143. https://doi.org/10.1016/j.fsi.2014.10.039.
- Zhang, C., Liu, G., Huang, M. 2014. Ribonucleotide reductase metallocofactor: assembly, maintenance and inhibition. Frontiers in biology, 9(2), 104–113. https://doi.org/10.1007/s11515-014-1302-6
- Zhang, Q. Y., Gui, J. F., 2018. Diversity, evolutionary contribution and ecological roles of aquatic viruses. Sci. China Life Sci. 61, 1486–1502. https://doi.org/10.1007/s11427-018-9414-7.
- Zhou, J., Wang, W.N., He, W.Y., Zheng, Y., Wang, L., Xin, Y., Liu, Y., Wang, A.L., 2010. Expression of HSP60 and HSP70 in white shrimp, Litopenaeus vannamei in response to bacterial challenge. J. Invertebr. Pathol. 103, 170–178. https://doi.org/10.1016/j.jip.2009.12.006.
No competing interests reported.
- Graficalabstract.jpg
Graphical Abstract
{kind=link}