The long chain polyunsaturated omega-3- fatty acids have been associated with many health effects due to their anti-inflammatory and cardio protective properties (Gogus & Smith, 2010; Poudyal et al., 2011). Especially eicosapentaenoic acid (EPA, C20:5) and docosahexaenoic acid (DHA, C22:6) play an essential role in the production of mediators, so called eicosanoids, that are important in regulating homeostatic functions (Cleland et al., 2003). By decreasing proinflammatory cytokines, lymphocytes proliferation and reactive oxygen species, the daily intake of DHA and EPA showed positive influence on the treatment and prevention of rheumatoid arthritis and coronary heart disease (Nielsen et al., 1992; Rennie et al., 2003; Volker et al., 2000). The recommended daily intake for a healthy human diet is a total of 250–500 mg DHA and EPA in a 1:1 ratio per day (Remize et al., 2021) A specific recommendation for DHA intake applies for young children and pregnant as well as lactating women (EFSA Panel on Dietetic Products & Allergies, 2012) .
DHA and EPA are synthesized in nature almost exclusively by marine microorganisms, especially microalgae, and are made available through fish and fish products, whereas terrestrial plants cannot produce these omega-3-fatty acids (Remize et al., 2021). Hence, microalgae are considered as primary producers of the marine food chain with regard to these fatty acids. Almost 90% of our omega-3-fatty acids supply comes from capture fisheries, often in form of fish oil supplements, as well as fishmeal and food fish (Tocher, 2015). According to the FAO (2018), sustainable fish stocks have reached a plateau, having declined from 90–66.9% between 1974 and 2015, while consumption of fish and marine products has increased significantly. Consequently, aquaculture filled the increasing demand. Since fish cannot produce DHA and EPA by themselves, aquaculture also relies on fish oil and fish meal from capture fisheries (Tocher et al., 2019). Increasing water temperature due to global warming also is thought to affect the composition of lipids in phytoplankton and microalgae by reducing the unsaturated fraction of lipids (Remize et al., 2021). In conclusion, the decline in sustainable fish stocks, the increase in aquaculture and a rising demand for fish products limit the global supply of polyunsaturated fatty acids from traditional sources and demonstrate the importance of finding alternative omega-3-fatty acid sources (FAO, 2018; Tocher et al., 2019).
One approach to solve the increasing gap, between DHA and EPA availability through the marine food chain, and the need of a growing world population for the essential fatty acids, is the heterotrophic production of algal oils by microalgae or microalgal-like microorganisms (Hamilton et al., 2015; Sijtsma & de Swaaf, 2004; Winwood, 2013). Using carbon sources such as glucose or acetic acid, heterotrophic marine microorganisms like Schizochytrium spp. and Crypthecodinium cohnii produce high amounts of PUFA’s and are currently used in commercial production (Barta et al., 2021; Sijtsma & de Swaaf, 2004; Tocher et al., 2019). Another approach involves transgenic oilseed plants that express the corresponding genes from microalgae, so that the vegetable oil can be enriched with DHA and EPA (Napier et al., 2015).
The use of photoautotrophically produced marine microalgae biomass with adequate DHA and EPA contents, as a dietary supplement, represents an attractive non-GMO omega-3-fatty acid alternative (Adarme-Vega et al., 2012). Phototrophic microalgae can accumulate high amount of lipids, including DHA and EPA and would be therefore a sustainable supply of these two health beneficial fatty acids (Ryckebosch et al., 2012). The use of saline or brackish water, the application of wastewater as fertilizer, no need of arable land and the capture of CO2 are further advantages for the photoautotrophic production of omega-3-fatty acids by microalgae (Draaisma et al., 2013; Polishchuk et al., 2015; Schenk et al., 2008; Wang et al., 2008).
Promising marine microalgae for the intracellular accumulation of DHA are e.g. Isochyrsis galbana or Tisochrysis lutea, accumulating 15–33 mgDHA gCDW−1 (Aussant et al., 2018; Guihéneuf & Stengel, 2013; Hu et al., 2018; Huerlimann et al., 2010). The marine golden-brown microalgal strains belong to the Haptophytes and are known for their use in aquaculture (Lin et al., 2007; Molina Grima et al., 1994). The genus Nannochloropsis (Eustigmatophyceae) is reported to accumulate high amounts of EPA with up to 50 mgEPA gCDW−1 (Aussant et al., 2018; Chua & Schenk, 2017; Gu et al., 2022; Hulatt et al., 2017; Polishchuk et al., 2015). Additionally the unicellular marine microalgae Nannochloropsis is already accepted as food (Chua & Schenk, 2017) and can be cultivated outdoors on a large scale (Rodolfi et al., 2009; San Pedro et al., 2014). Unfortunately, most marine microalgae strains do not accumulate both fatty acids, but the supply of DHA and EPA for human nutrition is recommended to be in a balanced ratio of 1:1. Consequently, production of microalgal biomass via a monoculture would not provide a nutritionally balanced food supplement for human nutrition (Ryckebosch et al., 2014; Tocher et al., 2019).
Co-cultivation of two marine microalgae may provide an opportunity to expand the product range via a photoautotrophic process. Due to the imbalanced DHA and EPA ratio of marine microalgae, co-culturing an EPA-producer together with a DHA-producer may be an option to obtain microalgal biomass enriched with balanced amounts of DHA and EPA.
A synthetic co-culture of two phylogenetically different marine phototrophic microalgal strains has been recently demonstrated using Tisochrysis lutea (Haptophyte) and Nannochloropsis oculata (Eustigmatophyceae) (Maglie et al., 2021): Mono- and co-cultures were conducted in dimly illuminated (60 µmol m− 2 s− 1) shake flasks applying a day/night rhythm. Under identical process conditions, N. oculata showed analogous growth behaviour in co-culture compared to that in monoculture, whereas T. lutea grew significantly worse in co-culture, so that at the end of the photoautotrophic batch processes in illuminated shake flasks, T. lutea accounted for only 17% of the total biomass. Despite the low percentage of T. lutea in co-culture, the omega-3-fatty acid DHA was detectable, resulting in an omega-3-fatty acid combination of EPA and DHA in a ratio of 13:1 in the harvested biomass.
Other studies showed, in principle, the feasibility of synthetic microalgae co-cultures (Rashid et al., 2019; Tejido-Nuñez et al., 2020). Tejido-Nuñez et al. (2020) reported a co-culture of the two microalgae strains Chlorella vulgaris and Tetradesmus obliquus at a pilot scale in open thin-layer cascade photobioreactors under realistic outdoor conditions: Using an inoculation ratio of 1:1, C. vulgaris accounted for only 10% of the final cells after a cultivation time of 29 days. A similar dominance effect of one species was reported by Zhao et al. (2014), when Chlorella sp. and Monoraphidium sp. were co-cultured in illuminated shake flasks. Nevertheless, the co-culture resulted in an enhanced lipid and biomass productivity. A co-culture of the two freshwater microalgae strains Ettlia sp. and Chlorella sp. achieved a higher biomass productivity compared to monocultures. Moreover, variation of the inoculation ratio resulted in an improved cell ratio of the two species in co-culture and further enhanced biomass productivity compared to 1:1 inoculated co-culture (Rashid et al., 2019).
Except the synthetic co-culture reported by Tejido-Nuñez et al. (2020), all studies were conducted at lab scale and at constant and dim light irradiance. Due to high operational and energy costs of closed indoor photobioreactors, most commercial microalgae processes are realized in open pond systems, normally under realistic outdoor climate conditions (Borowitzka, 1999). It has already been shown with photoautotrophic monocultures of microalgae that physical simulation of outdoor climate conditions (particularly dynamic incident light irradiation and temperature in day night cycles) is essential in lab-scale studies to enable the scale-up to outdoor photobioreactors (Pfaffinger et al., 2019; Wolf et al., 2021).
As the co-cultivation of the DHA-producing Tisochrysis lutea and the EPA-producing Nannochloropsis oculata on a dimly illuminated shake flasks scale was not successful with respect to the production of a microalgae biomass with EPA and DHA in a ratio of about 1:1 (Maglie et al., 2021), and the lab results may not easily be scalable to outdoor photobioreactors usually applied on a commercial scale for the production of marine microalgal biomass, we studied the photoautotrophic co-cultivation of the DHA-producing Tisochrysis lutea with the alternative EPA-producer Microchloropsis salina applying realistic physical climate simulation in the lab-scale photobioreactors. Formerly, strains from the genus Microchloropsis belonged to the genus Nannochloropsis and therefore show strong genetically similarity (Fawley et al., 2015). Under nitrogen replete conditions, good EPA production such as 16 mgEPA gCDW−1 were reported for Microchloropsis salina (Gu et al., 2022; Schädler et al., 2019).
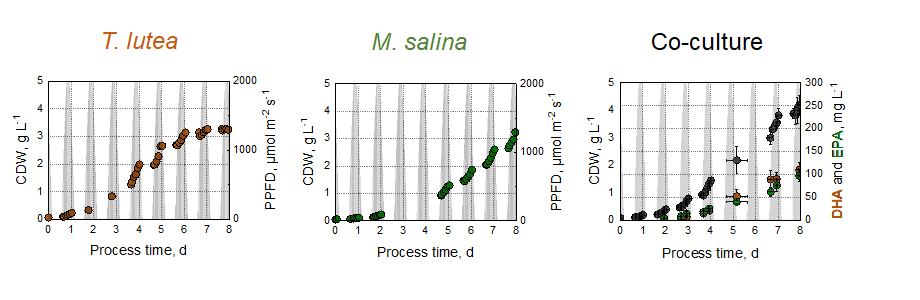
In this work, we report on co-culture studies with the two marine phylogenetically different microalgal strains, T. lutea and M. salina, in artificial sea water for the photoautotrophic production of DHA and EPA balanced microalgae biomass in 1.8 L flat-plate gas-lift photobioreactors applying a dynamic climate simulation of a repeated sunny summer day in Australia. The temperatures ranged between 15–30°C and the incident photon flux densities were varied between 0-2000 µmol m− 2 s− 1 in order to investigate biomass growth and omega-3-fatty acid production of both strains in monoculture and in co-cultures with varying inoculation ratios in the batch processes.
{kind=link}