Flower colors of 'Sunset Babylon Eyes' at different developmental stages
We collected materials at five stages depending on the progress of anthesis (Fig. 1a). At S1 stage, petals were yellow-green without blotches. Cerise blotches appeared at the base of the light yellow petals at S2 stage. At S3 stage, blotches grew and turned rose-red while non-blotch parts were butter yellow. Blotches grew continuously and their color turned crimson, and the non-blotch parts became bright yellow at S4 stage. At S5 stage, non-blotch parts were yellow while blotches turned dark red.
To precisely evaluate the color of rose flowers, color parameters L*, a*, and b* of CIEL*a*b* color system of petals were measured. Significant differences were observed in color parameters among the blotch and non-blotch parts at different stages (Fig. 1b). Lightness (L*) value of the blotch and non-blotch parts can be divided into three groups: S1, S2-S3 and S4-S5. From S2 to S5 stages, L* value of the non-blotch parts increased, while that of the blotch parts decreased. Also, L* value of the non-blotch parts at S4 and S5 stages was even higher than that of the petals at S1 stage. Parameter a* represents green and red color from negative value to positive value, and parameter b* represents blue and yellow color from negative value to positive value. The a* value and b* value of the non-blotch parts were nearly close at S2 and S3 stages, and they increased significantly at S4 stage, while they decreased again at S5 stage. From S2 to S5 stages, the a* value of the blotch parts showed a fluctuation trend of 'increase-decrease-increase', and so the overall trend was from green to red, then to green, and finally turned to red. The b* value of the blotch parts decreased from S2 to S4 stages, and slightly increased at S5 stage, thus the overall trend increasing from yellow to blue. The overlap of a* value and b* value of the blotch parts at S5 stage showed that the blotch parts are so red and purple.
Identification and quantification of flavonoids in petals of 'Sunset Babylon Eyes'
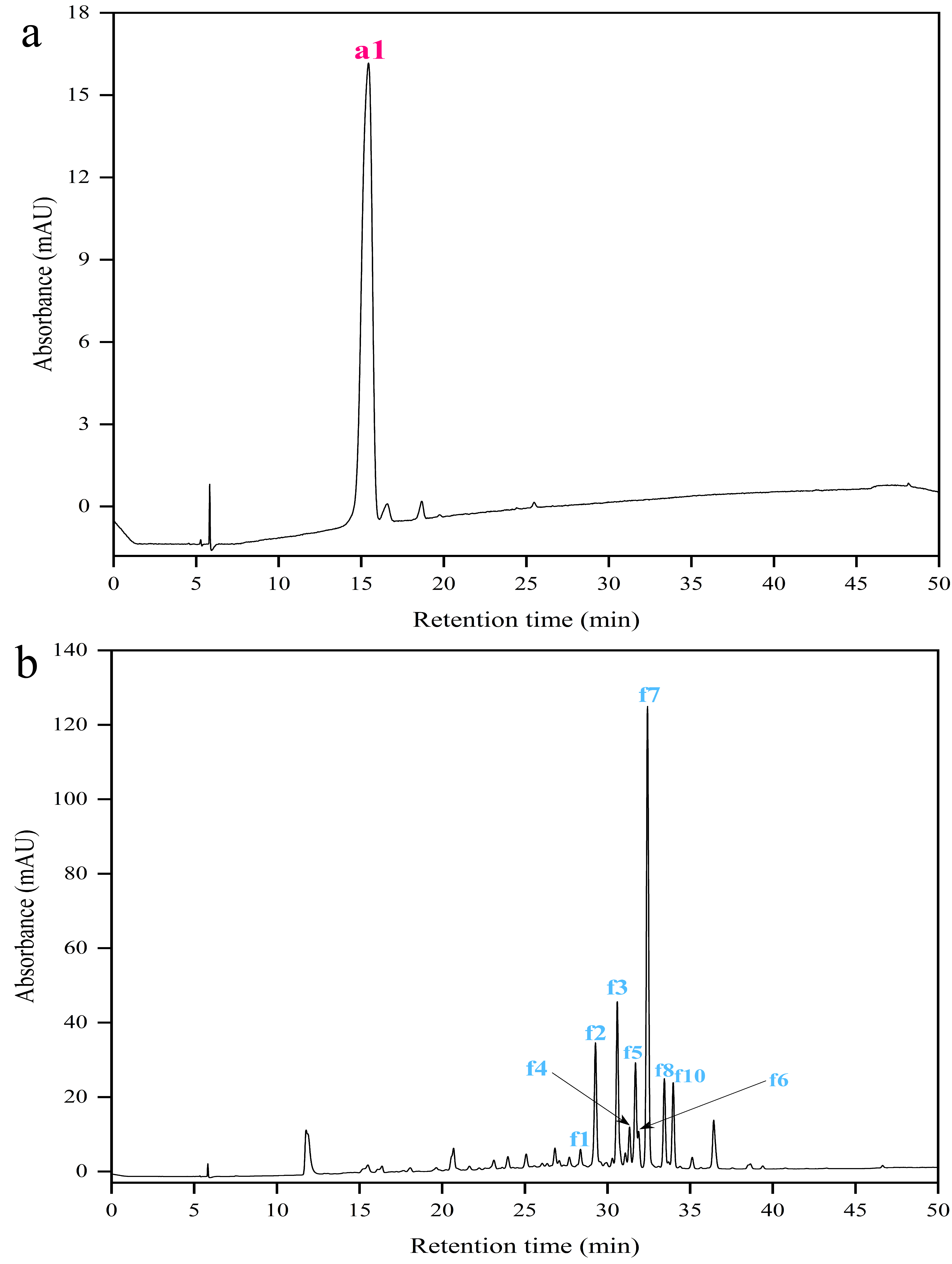
Under the wavelength of 525 nm, the extraction samples of petal powder at different developmental stages of 'Sunset Babylon Eyes' were detected by HPLC-DAD. The color blotch from S2 to S5 stages were detected and only one anthocyanin: cyanidin 3,5-O-diglucoside (Cy3G5G) was found (Table 1, Supplementary Fig. 2a, Supplementary Table S1). The standard curves were plotted with different concentrations of Cy3G5G standard products. The concentration (mg/mL) was used as the abscissa and the peak area was used as the ordinate to obtain the corresponding linear regression equation y = 22887x-96.484 (R2 = 0.9987). Anthocyanins were not detected at S1 stage and in the non-blotch parts from S2 to S5 stages. The content of Cy3G5G is also the total contents of anthocyanin (TA) in 'Sunset Babylon Eyes' petals (Fig. 1c) The content of Cy3G5G was very low in the blotch parts at both of S2 and S3 stages. TA in the blotch part at S4 stage was the highest, which was about 7.4 times higher than that of the blotch parts at S2 stage. And TA in the blotch part at S5 stage was close to but slightly lower than that at S4 stage.
Table 1
Identification of flavonoids.
| Peak no. | Retention time (min) | λvis−max (nm) | λvis−acyl (nm) | ESI−MS−(m/z) | Aglycone | Main identified molecule | References | Standard |
| a1 | 15.708 | 511 | 277 | 611, 449, 287 | Cyanidin | Cyanidin 3,5-O-diglucoside | / | Cyanidin 3,5-O-diglucoside |
| f1 | 28.061 | 328 | 257 | 433, 301 | Quercetin | Quercetin 3-O-α-L-arabinofuranoside | [36, 38, 72] | / |
| f2 | 29.021 | 340 | 257 | 593, 431, 285 | Kaempferol | Kaempferol 3-O-rhamnosyl-7-O-glucoside | [39] | Kaempferol 3-O-rhamnoside-7-O-glucoside |
| f3 | 29.984 | 359 | 254 | 609, 447, 301 | Quercetin | Quercetin 3-O-rutinoside | [35] | Quercetin 3-O-rutinoside |
| f4 | 30.328 | 347 | 265 | 447, 285 | Kaempferol | Kaempferol 3-O-glucoside | [35, 38] | Kaempferol 3-O-glucoside |
| f5 | 30.816 | 347 | 256 | 447, 301 | Quercetin | Quercetin 3-O-rhamnoside | [72] | Quercetin 3-O-rhamnoside |
| f6 | 31.118 | 360 | 257 | 463, 301 | Quercetin | Quercetin 3-O-galactoside | [35, 38, 72] | Quercetin 3-O-galactoside |
| f7 | 31.621 | 347 | 257 | 599, 447, 285 | Kaempferol | Kaempferol 3-O-(galloyl)-glucuronide or kaempferol 7-O-(galloyl)-glucuronide | [39] | / |
| f8 | 32.164 | 359 | 265 | 447, 285 | Kaempferol | Kaempferol 7-O-glucoside | [35, 38] | Kaempferol 7-O-glucoside |
| f9 | 33.168 | 347 | 265 | 417, 285 | Kaempferol | Kaempferol 3-O-arabinoside | [36, 72] | Kaempferol 3-O-arabinoside |
| f10 | 33.734 | 304 | 265 | 431, 285 | Kaempferol | Kaempferol 3-O-α-D-rhamnoside | [35, 36, 38, 72] | Kaempferol 3-O-α-D-rhamnoside |
| f11 | 46.593 | / | / | 285 | Kaempferol | Kaempferol | [39] | Kaempferol |
Eleven flavonol glycosides were detected and identified by HPLC-DAD, and no flavonoid glycosides (Supplementary Fig. 2b; Table 1) were detected. Kaempferol (f11) was only detected at S1 stage. Kaempferol 3-O-α-D-rhamnoside (f10) was not detected at S1 and S2 stages, but the compound began to accumulate at S3 stage. Among the flavonol glycosides, f7, which was characterized as kaempferol 3 -O-(galloyl)-glucuronide or kaempferol 7 -O-(galloyl)-glucuronide, was the predominant component in the blotch parts at S1, S2 and S3 stages, and accumulated from S1 to S4 stages, and no longer accumulated at S5 stage. However, f8 (kaempferol 7 -O-glucoside) and f10 (kaempferol 3 -O-α-D-rhamnoside) were the predominant flavonols in the petals of non-blotch parts at S3, S4 and S5 stages (Supplementary Table S1). Therefore, kaempferol glycosides may play an important role in the coloration of the non-spot parts of 'Sunset Babylon Eyes'. At S1 and S2 stages, f7 (kaempferol 3-O-(galloyl)-glucuronide or kaempferol 7-O-(galloyl)-glucuronide) was the key compound for petals to show yellow. From S3 to S5 stages, f8 (kaempferol 7 -O-glucoside) played a major role in coloration.
The standard curve was drawn with different concentrations of quercetin 3-O-galactoside (Qu3Gal) standard. The linear regression equation y = 46796x + 10.026 (R2 = 0.9996) was obtained with concentration (mg/mL) as abscissa and peak area as ordinate. The total contents of flavones and flavonols (TF) at S5 stage was the lowest, followed by that at S1 stage. While TF in the non-blotch parts were always higher than that in the blotch parts. Especially in the non-blotch part at S3 stage, TF reached the maximum (Fig. 1c). The content of total kaempferol and its derivatives (Total Km) was always higher than that of total quercetin and its derivatives (Total Qu) at all surveyed stages. From S1 to S3 stages, Total Km and Total Qu increased. The contents of total flavones and flavonols (TF) were similar between S4 and S5 stages, but Total Qu at S5 stage was higher than that at S4 stage. Total Qu of the non-blotch parts was the lowest at S3 stage, and there was little difference in other stages. Total Km increased gradually from S1 to S3 stages, and reached the peak at S3 stage, and subsequently decreased gradually from S3 to S5 stages. However, Total Km of the non-blotch part at S5 stage was higher than that of the non-blotch part at S2 stage (Fig. 1d). Consequently, the yellow non-blotch parts of 'Sunset Babylon Eyes' may be related to the synthesis of flavonoids.
Identification and quantification of carotenoids in petals of 'Sunset Babylon Eyes'
Twelve carotenoids (Supplementary Table S1, Supplementary Table S2) were detected and identified by HPLC-DAD. The standard curve was drawn with different concentrations of 13/13’Z-Neoxanthin standard. The linear regression equation y = 288930x-614.38 (R2 = 0.9999) was obtained with concentration (mg/mL) as abscissa and peak area as ordinate. Petals at S1, S2 and S3 stages contained only two kinds of carotenoids ((all-E)-Lutein (c7) and (all-E) -β-Carotene (c12)), and the content of c7 was always higher than that of c12. At S4 and S5 stages, the content of c7 decreased and the distribution of each component was relatively average, but the content of c12 was the highest. The total contents of carotenoids (TC) at S1 stage were slightly higher than that at S2 and S3 stages. The types and the total contents of carotenoids began to surge from S4 stage, and both of them reached the maximum at S5 stage. At S4 and S5 stages, the carotenoids composition were not the exactly same in the blotch parts and in the non-blotch parts (Supplementary Table S1). Petals in the non-blotch part at S5 stage contained the most kind of carotenoids and the total contents of carotenoids (TC) at S5 stage was the highest.
Thus, TC can be divided into a group of S1, S2-S3 and S4-S5, which can be completely corresponding to the brightness of petals (L*) (Fig. 1c). Therefore, carotenoids may mainly have impact on the brightness of 'Sunset Babylon Eyes' petals, which made the petals show a brighter color. In terms of pigmentation in the blotch and non-blotch parts, therefore, candidate differentially expressed genes (DEGs) from the flavonoid biosynthesis pathway but not the carotenoids biosysnthesis pathway would be searched for subsequent experiment.
Transcriptome analysis of 'Sunset Babylon Eyes'
To study the molecular basis of petal blotch formation in rose, library preparation and RNA-seq were performed on non-blotch and blotch parts at five stages. After filtering these raw reads, the clean reads were obtained, which ranged from 36716138 to 43862630 (Supplementary Table S3). The Q30 value of all 27 libraries were more than 91%. Between 81.28% and 83.30% of the sequenced reads could be aligned to the rose reference genome (Supplementary Table S3).
Comparisons of the DEGs between non-blotch and blotch parts at different developmental stages
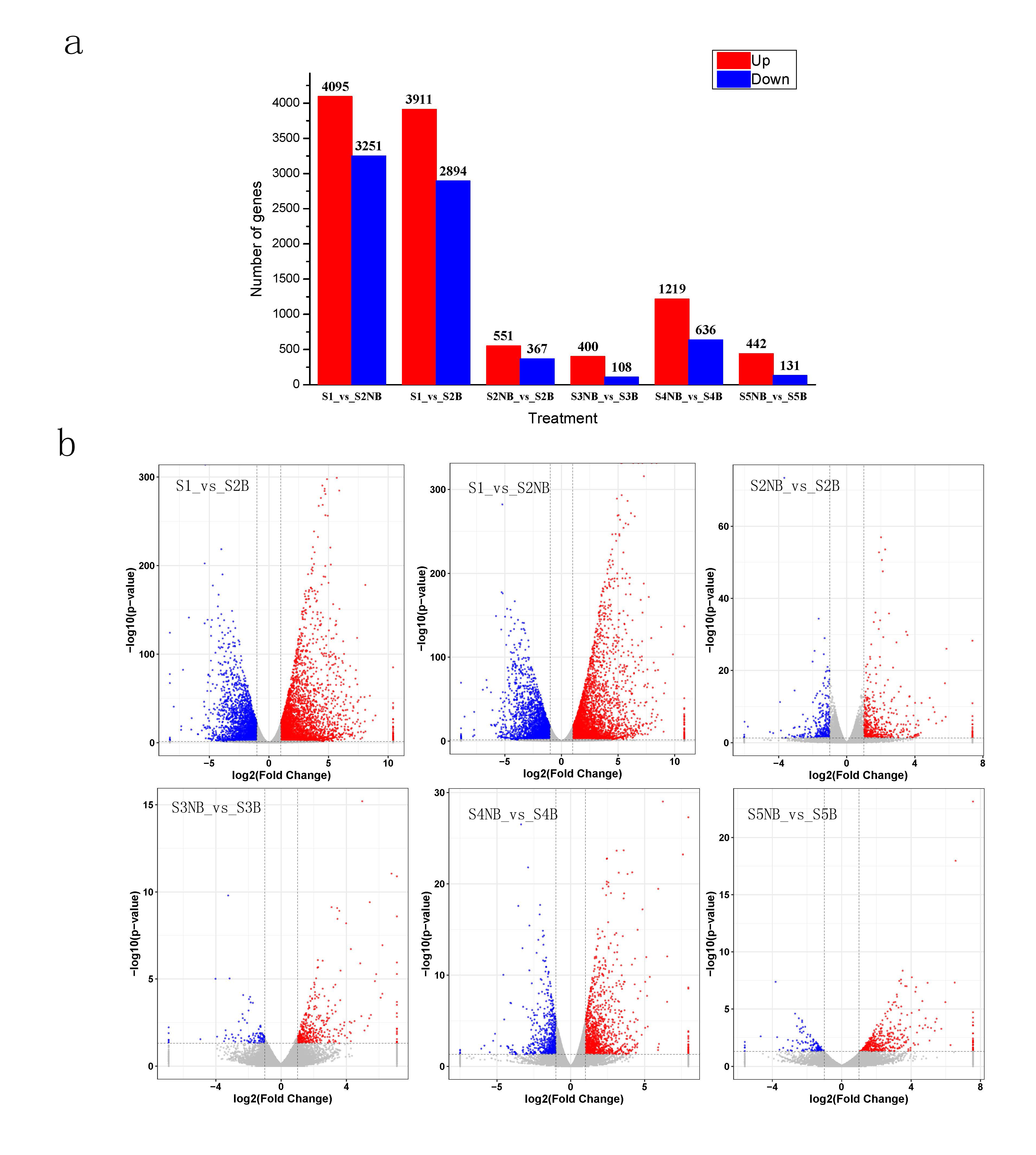
To analyze the DEGs between non-blotch and blotch parts at different developmental stages, comparisons were conducted between six groups. The DEGs were filtrated according to an expression level |log2(FC) | > 1 and p-value < 0.05 in each pairwise comparison. Upregulated DEGs and downregulated DEGs were counted, as shown in Supplementary Fig. 3. The number of DEGs was much higher at the S1vs S2B and S1vs S2NB comparisons than other four comparisons, indicating S2 stage, which was the time the petal blotch appeared, was an important stage for the formation of petal blotch. From S2 to S5 stages, the number of DEGs reached a peak at the S4NB vs. S4B comparison (1855 upregulated/downregulated genes) and was lowest at the S3NB vs. S3B comparison (508 upregulated/downregulated genes), illustrating that there are dramatic changes between non-blotch and blotch parts at S4 stage and opposite at S3 stage. Considering that anthocyanins only accumulated in blotch parts, the results were found that there were only 9 upregulated DEGs and 4 downregulated DEGs in S1vsS2B, S2NBvsS2B, S3NBvsS3B, S4NBvsS4B and S5NBvsS5B datasets (Supplementary Fig. 4), and all DEGs were not related to anthocyanin synthesis pathway. These results indicated that different genes affected the anthocyanin accumulation at different petal developmental stages.
GO functional enrichment and KEGG pathway enrichment analysis of DEGs
We analyzed the GO and KEGG pathways to determine the biological functions of DEGs. 16,367(41.26%) and 13,695(34.52%) genes were annotated to GO and KEGG databases. The GO annotation system consisted of three major branches: biological process, molecular function, and cellular component. Phosphotransferase activity, cinnamyl − alcohol dehydrogenase activity, xyloglucosyl transferase activity, sequence − specific DNA binding, secondary active sulfate transmembrane transporter, and isoleucine − tRNA ligase activity were the most significant enrichment GO terms under the molecular function category among six comparisons, respectively. Meanwhile, under the biological process category, DNA packaging, defense response, xyloglucan metabolic process, auxin metabolic process, sulfate transport, and response to biotic stimulus were the most significant enrichment GO terms among six comparisons, respectively (Supplementary Fig. 5).
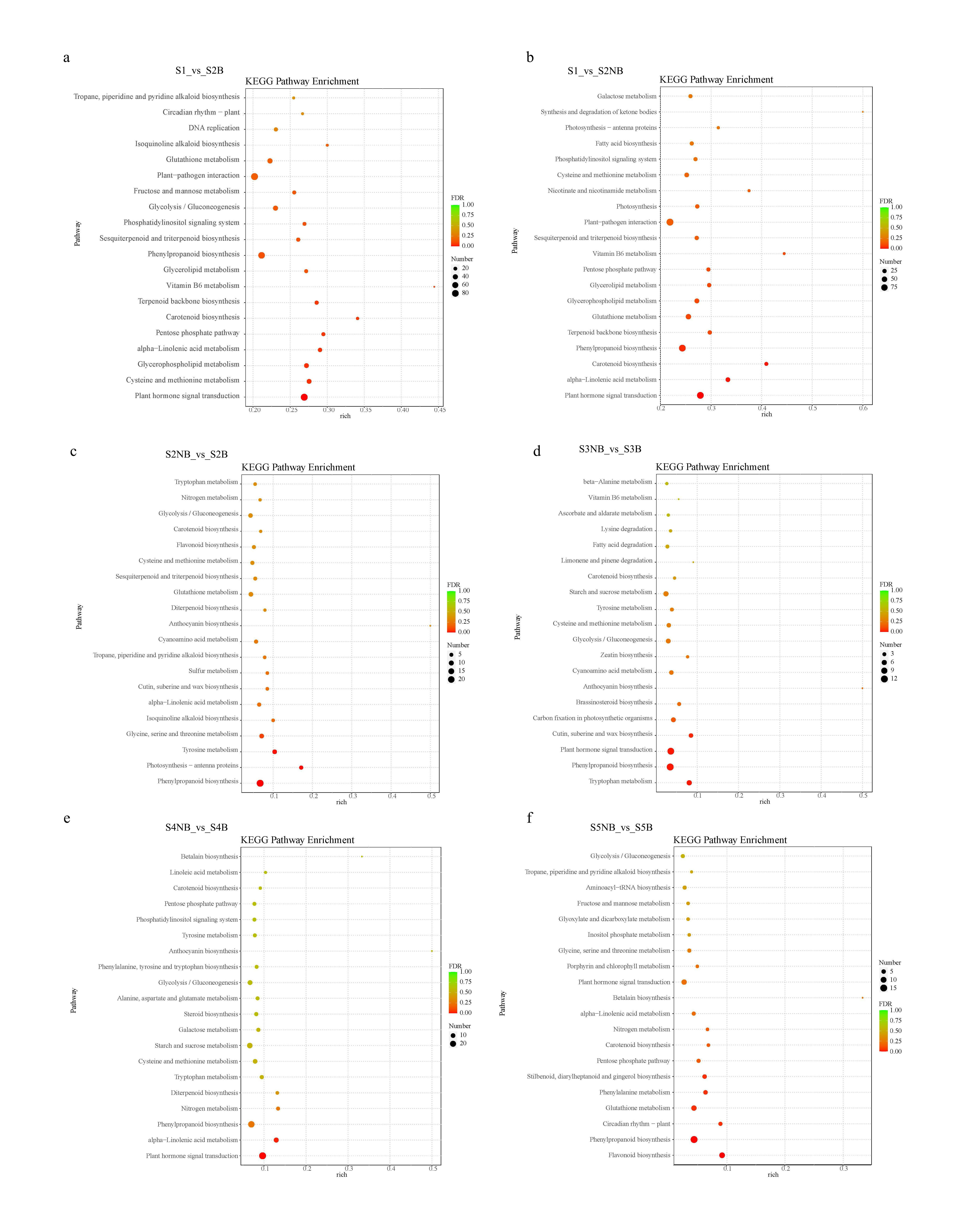
In the KEGG signal enrichment pathway, phenylpropanoid pathway and carotenoid biosynthesis were enriched at each comparison. Phenylpropanoid pathway was the upstream pathway of flavonoid biosynthesis and anthocyanin synthesis. In the S2NB vs. S2B, S3NB vs. S3B and S4NB vs. S4B comparisons, anthocyanin synthesis pathway was enriched. Meanwhile, flavonoid biosynthesis pathway was enriched in S2NB vs. S2B and S5NB vs. S3B comparisons (Supplementary Fig. 6).
Mining of enzyme genes in the transcriptome of 'Sunset Babylon Eyes'
To investigate the pathways of pigments synthesis in the blotch and non-blotch parts of petals, we analyzed the genes involved in anthocyanin and flavonol biosynthesis pathways in Rosa 'Sunset Babylon Eyes'. After searching all these genes in the functional annotations, a total of 143 genes involved in the above pathways (Table 2) were found considering the change of the contents of pigments, and the transcriptional profiles of the genes involved in anthocyanins biosynthetic pathway between blotch and non-blotch parts from S1 to S4 stages and genes involved in flavonol biosynthesis pathway between blotch and non-blotch parts from S1 to S3 stages.
Table 2
Candidate unigenes involved in anthocyanin and flavonol biosynthesis in rose.
| Function | Gene name | Gene abbrevia-tion | Total No. | DEG No. |
| Flavonoid synthesis pathway | Chalcone synthase | CHS | 7 | 3 |
| Chalcone isomerase | CHI | 7 | 5 |
| Flavanone 3-hydroxylase | F3H | 4 | 2 |
| Flavonoid 3’-hydroxylase | F3’H | 15 | 1 |
| Flavonoid 3′,5′-hydroxylase | F3’5’H | 5 | 0 |
| Dihydroflavonol 4-reductase | DFR | 6 | 2 |
| Anthocyanin synthesis pathway | Anthocyanidin synthase | ANS | 7 | 2 |
| UDP-glucose: anthocyanin 3-glucosylltransferase | A3GT | 11 | 3 |
| Anthocyanidin 3-O-glucoside 2″-O-glucosyltransferase Anthocyanidin 3-O-glucoside 6″-O-glucosyltransferase | A3GGT | 7 | 0 |
| Anthocyanidin 5, 3-O-glucosyltransferase | A5, 3GT | 3 | 0 |
| Flavone and flavonol biosynthesis pathway | flavonoid 3-glucosyltransferase | F3GT | 4 | 0 |
| UDP-glucose flavonoid 3-O-glucosyltransferase | 52 | 6 |
| UDP-glucose: flavonol 3-glucosyltransferase | 3 | 0 |
| Flavonol synthase | FLS | 12 | 4 |
| Transcription factors | AP2/B3 transcription factor family protein | AP2 | 13 | 1 |
| basic helix-loop-helix (bHLH) DNA-binding superfamily protein | bHLH | 63 | 7 |
| Dof-type zinc finger DNA-binding family protein | DOF | 13 | 3 |
| myb domain protein | MYB | 85 | 9 |
| NAC domain containing protein | NAC | 119 | 8 |
| TCP domain protein | TCP | 9 | 3 |
| Transducin/WD40 repeat-like superfamily protein | WD40 | 235 | 6 |
| WRKY DNA-binding protein | WRKY | 55 | 6 |
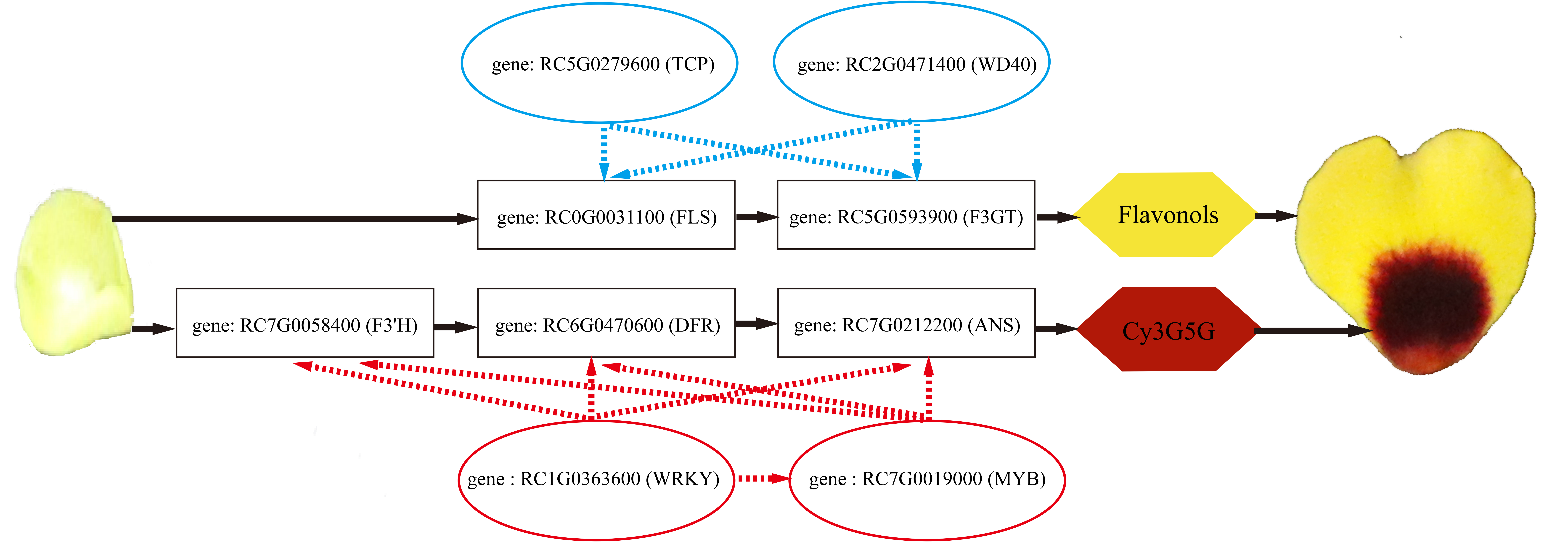
A total of 28 key enzyme genes were found, which contained upstream genes (CHS, CHI, etc.) and downstream genes (DFR, ANS, etc.) (Fig. 2, Table 2). As the upstream genes of anthocyanin and flavonol biosynthesis pathways, the majority of CHS, CHI and F3H genes expressed higher at early stages (Fig. 2). Among them, two enzyme genes, RC1G0025000 (chalcone synthase, CHS) and RC1G0494500 (chalcone isomerase, CHI), both had the highest expression level at S1 stage and expressed higher in blotch parts from S2 to S3 stages. RC2G0136900 (flavanone 3-hydroxylase, F3H) has the highest expression level at S5 stage, except that the expression level in the blotch part at S2 stage was higher, from S3 to S5 stages, its expression in the non-blotch parts was lower than or equal to that in the blotch parts.
FLS and F3GT were key enzyme genes for the biosynthesis and modification of flavonol compounds, thus genes which expressed higher in the non-blotch parts from S1 to S3 stages were got more attention. Among them, the expression level of RC0G0031100 (Flavonol synthase, FLS) was the highest in the non-blotch part at S2 stage. In addition, RC5G0593900 (UDP-glucose: anthocyanin 3-glucosylltransferase, A3GT), whose expression level was higher in blotch parts from S1 to S3 stages, could also be the key gene for the modification of flavonol compounds.
As the downstream genes of anthocyanin synthesis pathway, the majority of F3’H, DFR, and ANS genes expressed higher in the blotch parts from S2 to S4 stages. The results demonstrated that upstream genes (CHS, CHI) of the synthesis of anthocyanin and flavonol expressed highest at early stages. The downstream genes (F3’H, DFR, ANS) of anthocyanin synthesis pathway expressed higher in the blotch parts from S2 to S4 stages, and this would accumulate the amount of cyanidin in the blotch parts. The downstream gene (FLS, F3GT) of flavonol synthesis pathway expressed higher in non-blotch part at S2 stage, which may lead to the accumulation of flavonol compounds in non-blotch parts.
Mining of regulatory genes in the transcriptome of 'Sunset Babylon Eyes'
Some transcription factor (TF) families including MYB, bHLH, AP2, DOF, NAC, TCP, WD40 and WRKY play important roles in color formation via anthocyanins biosynthesis by regulating the expression of key enzyme genes. We therefore analyzed the expression pattern of 592 genes belonging to these TF families in blotch and non-blotch parts (Table 2) and 43 DEGs were found (Fig. 3). Of 43 genes, there were 1 AP2 gene, 7 bHLH genes,13 DOF genes,9 MYB genes,8 NAC genes, 3 TCP genes, 6 WD40 genes and 6 WRKY genes.
Among them, RC5G0279600 (TCP) and RC2G0471400 (WD40) had the highest expression level at S1 stage and the expression level in all the blotch parts were lower than or equal to those in the non-blotch parts. The flower color change of blotch and non-blotch parts of 'Sunset Babylon Eyes' may be related to the decrease of the expression of these two regulatory genes. 8 regulatory genes were up-regulated in blotch parts (Fig. 3). RC7G0019000 (MYB) was an up-regulated gene in blotch parts and expressed highest at S4 stage. However, as up-regulated genes in blotch parts, RC1G0363600 (WRKY), RC2G0406300 (AP2), RC5G0166100 (WRKY), RC1G0093800 (NAC), RC2G0093100 (MYB), RC0G0030000 (DOF), and RC0G0179800 (NAC) expressed at the highest level in the blotch part at S2 stage. Seven bHLH genes highly expressed at S1 stage or in the non-blotch parts, so they may not be the key transcription factors that mostly directly affects the blotch pigmentation.
Collectively, RC5G0279600 (TCP) and RC2G0471400 (WD40) may be negative regulatory genes for blotch formation. RC7G0019000 (MYB), RC1G0363600 (WRKY), RC2G0406300 (AP2), RC5G0166100 (WRKY), RC1G0093800 (NAC), RC2G0093100 (MYB), RC0G0030000 (DOF), and RC0G0179800 (NAC) may be positive regulatory genes for blotch formation. RC7G0019000 (MYB) may be involved in the regulation of the late stages of flower color synthesis in 'Sunset Babylon Eyes', while other 7 genes might be involved in the regulation of the early stages of 'Sunset Babylon Eyes' flower color synthesis.
Quantitative real-time PCR (qRT-PCR) analysis of candidate enzyme genes and regulatory genes
qRT-PCR analysis was performed on 9 candidate enzyme genes and 10 candidate regulatory genes, and it was found that the fitting rate of expression trends with transcriptome data in each stage reached 89.47%. According to the qRT-PCR results, RC1G0025000 (CHS), RC1G0494500 (CHI), RC2G0136900 (F3H) may be involved in the initiation of 'Sunset Babylon Eyes' blotch, and RC7G0563900 (ANS) may be related to the deepening of 'Sunset Babylon Eyes' blotch in the later stages, but the relative expression level of these four enzyme genes was not much different from those of the non-blotch parts. Two enzyme genes, RC0G0031100 (FLS) and RC5G0593900 (A3GT), significantly highly expressed in the non-blotch parts at S2 and S3 stages, respectively, and they may be involved in the synthesis of flavonol glycosides in the non-blotch parts. Three enzyme genes, RC7G0058400 (F3'H), RC6G0470600 (DFR) and RC7G0212200 (ANS), all significantly highly expressed in the blotch parts at S2 and S4 stages, which should be the key enzyme genes for the early pigmentation and color deepening of blotch in the later stages (Fig. 4).
RC2G0093100 (MYB) significantly highly expressed in the blotch part at S2 stage, but also highly expressed in the non-blotch part at S3 stage. RC2G0406300 (AP2) significantly highly expressed in the blotch part at S2 stage, however, at S4 stage, its relative expression level of the blotch part was lower than that of the non-blotch part. These two regulatory genes may be involved in the regulation of the early pigmentation in blotch parts, or also be related to the regulation of the late pigmentation in non-blotch parts at the same time. Six regulatory genes, including RC7G0019000 (MYB), RC1G0363600 (WRKY), RC5G0166100 (WRKY), RC0G0179800 (NAC), RC1G0093800 (NAC), and RC0G0030000 (DOF), from S2 to S4 stages, the relative expression level of the blotch parts was all higher than that of the non-blotch parts. It was particularly worth mentioning that RC7G0019000 (MYB) and RC1G0363600 (WRKY) significantly highly expressed at S2 and S4 stages, and both of them may be the key regulatory genes for the early pigmentation and color deepening of blotch in the later stages. While the relative expression level of the rest two genes, RC5G0279600 (TCP) and RC2G0471400 (WD40), in blotch parts from S2 to S4 stages were lower than those in non-blotch parts, in that case, these two genes may negatively regulate the pigmentation of blotch (Fig. 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}