Overview of Sequencing Data
To understand the expression characteristics of circRNA in porcine muscle at different developmental stages, we performed ribosomal RNA-depleted RNA-seq on muscle sampled at three time points. Firstly, we constructed nine non-coding RNA libraries named JFW_1d_1, JFW_1d_2, JFW_1d_3, JFW_90d_1, JFW_90d_2, JFW_90d_3, JFW_180d_1, JFW_180d_2, and JFW_180d_3. Secondly, we purified and sequenced the RNA using the Illumina paired-end RNA-seq method. For these libraries, 908,657,602 raw reads were obtained from the platform, and after filtering, a final dataset of 902,652,752 clean reads was obtained (Table 1). The JFW_180d_3 group contained the greatest number of reads (106,567,824), and the JFW_1d_1 group had the least (91,942,766). The Q30 quality threshold (Phred quality score >30) was met for 95.2% of the data. Taken together, these results confirmed that the sample and sequencing data generated in this study were of good quality and were reliable for subsequent bioinformatic analysis.
Table 1. Overview of RNA-seq results for each sample.
|
Sample
name
|
Raw
reads
|
Clean
reads
|
Raw_bases
(G)
|
Clean_bases
(G)
|
Error rate
(%)
|
Q20
(%)
|
Q30
(%)
|
|
JFW_1d_1
|
92,655,706
|
91,942,766
|
13.9
|
13.79
|
0.02
|
98.62
|
95.72
|
|
JFW_1d_2
|
107,050,232
|
106,444,968
|
16.06
|
15.97
|
0.02
|
98.64
|
95.78
|
|
JFW_1d_3
|
104,392,040
|
103,814,406
|
15.66
|
15.57
|
0.02
|
98.58
|
95.45
|
|
JFW_90d_1
|
96,202,328
|
95,550,798
|
14.43
|
14.33
|
0.03
|
98.08
|
94.2
|
|
JFW_90d_2
|
106,311,518
|
105,563,842
|
15.95
|
15.83
|
0.02
|
98.56
|
95.75
|
|
JFW_90d_3
|
95,622,436
|
94,984,938
|
14.34
|
14.25
|
0.02
|
98.67
|
95.89
|
|
JFW_180d_1
|
94,853,354
|
94,375,978
|
14.23
|
14.16
|
0.03
|
97.81
|
93.78
|
|
JFW_180d_2
|
104,189,042
|
103,407,232
|
15.63
|
15.51
|
0.02
|
98.59
|
95.54
|
|
JFW_180d_3
|
107,380,946
|
106,567,824
|
16.11
|
15.99
|
0.02
|
98.40
|
95.04
|
|
Total
|
908,657,602
|
902,652,752
|
15.15
|
15.04
|
0.02
|
98.44
|
95.24
|
Characterization of circRNAs in Porcine Skeletal Muscle
After mapping the clean data to the pig genome (Sscrofa11.1) using the find_circ and CIRI software, we identified a total of 16,990 highly credible circRNAs from the nine libraries (summarized in more detail in Supplementary Table S1). In addition, we mapped the chromosomal distribution of these circRNAs, finding them to be transcribed from all chromosomes but with an uneven distribution (Figure 1A). In agreement with other earlier work[39], we observed that the highest number of circRNAs originated from chromosomes 1, 6, and 13 in this study (Figure 1A). Furthermore, since circRNAs are primarily derived from annotated exons[40], we also found that >90% identified circRNAs in this study contained exon sequences (Figure 1B). The length distribution of the predicted circRNAs showed that most of them were less than 1,000 nt in length (Figure 1C), which was also consistent with previous reports[41].
Differential Expression of circRNAs
To better understand the differences in circRNA expression patterns in pig muscle at 1, 90, and 180 days, we used DESeq software to identify DE circRNAs between sample pairs (i.e., JFW_90d vs. JFW_1d, JFW_180d vs. JFW_1d, and JFW_180d vs. JFW_90d). We also performed hierarchical clustering analysis of the DE circRNAs. This revealed distinct differential expression patterns for the identified circRNAs between the three time points in muscle, with JFW_1d showing significant differences compared to the other two groups (Figure 2A). In total, 584 DE circRNAs were detected using the above pairwise comparisons. The number of differentially expressed genes was 477, 63, and 255 for the JFW_180d vs. JFW_1d, JFW_180d vs. JFW_90d, and JFW_90d vs. JFW_1d comparisons, respectively (Figure 2B). We further constructed a Venn diagram of the identified DE circRNAs in the three pairwise comparisons to find the shared circRNAs (Figure 2C). It revealed that only two circRNAs (novel_circ_0009188 and novel_circ_0018581) were consistently differentially expressed in all comparisons (Figure 2C). One of these, novel_circ_0009188, is generated by circularization of two exons of tubulin tyrosine ligase-like 7 (TTLL7) and is 311 nt in length. The other, novel_circ_0018581, arises through circularization of two exons of glutamate decarboxylase-like protein 1 (GADL1), and has a length of 608 nt. The expression levels of these two circRNAs increased significantly with age in Jinfen White pigs, across the three growth stages.
Validation of DE circRNAs at Different Developmental Stages of Muscle
To validate the reliability of temporal circRNA expression profiles derived from RNA-seq data, divergent primers were designed for four randomly selected DE circRNAs, and the expression levels of that were quantified by qRT-PCR (Figure 3A). The results were consistent with the expression patterns of these circRNAs obtained through RNA-seq data. In addition, the expected size PCR products were obtained by amplification using cDNA as the PCR template. The post-splice sites were verified by Sanger sequencing (Figure 3B). These results suggested that the identified circRNAs in this study are credible.
Functional Enrichment Analysis of DE circRNA Host Genes
Previous studies have shown that circRNAs regulate host gene transcription by competing with linear pre-mRNA splicing[42,43]. To explore the potential functions of the identified DE circRNAs in the development of skeletal muscle, we performed GO and KEGG pathway enrichment analyses of their host genes. Notably, we found that these host genes mainly have known roles in muscle biology, including muscle contraction, muscle organ development, muscle system processes, and muscle structure development (Figure 4A). The host genes CHD2, SMAD3, BMPR1A, HOMER1, DMD, and MYL2 are involved in muscle organ development; HOMER1, DMD, and ATP11A are involved in myotube differentiation; and NFATC1, BMPR1A, and PDCD4 are involved in the negative regulation of muscle cell differentiation.The KEGG pathway enrichment analysis showed that the DE circRNA host genes were enriched in 20 pathways, including skeletal muscle fiber-related signaling pathways, and the cAMP and AMPK signaling pathways et al (Figure 4B). PDE4B, NFATC1, PLCE1, ATP2B4, PPP1R12A, and CAMK2B were enriched in the cAMP signaling pathway, and EEF2K, FBP2, PPP2CB, PFKFB1, and PRKAA2 were enriched in AMPK signal pathway. The circRNAs generated from these host genes might act as promoters or repressors during the development of porcine skeletal muscle, thereby providing candidate genes for future exploration of the regulatory functions of these genes and the circRNAs derived from them.
Construction of a potential circRNA-miRNA-mRNA regulatory network
CircRNAs often act as miRNA molecular sponges to indirectly regulate the expression of miRNA target genes, thereby meaning that they participate in the related biological processes[44]. Therefore, we integrated miRNA and mRNA library data (The sequencing results obtained at the same time, the accession number is consistent with the circRNA) with the results from the present study to constructed a ceRNA network and identified key circRNAs associated with pork quality and regulation of cell proliferation and differentiation. This ceRNA network contained 229 circRNAs, 55 miRNAs and 143 mRNAs (Supplementary Table S2). Because the ceRNA network are too large, we selected the top 20 DE circRNAs with the most obvious up- or down-regulation profiles to present two ceRNA network maps (Figure 5). In ceRNA network, novel_circ_0018595 (hereafter, circ_0018595) is predicted to function as a sponge for ssc-miR-1343, which binds to the mRNAs transcribed from the PGM1, TNNT3, and IFNAR2 genes (Figure 5A).
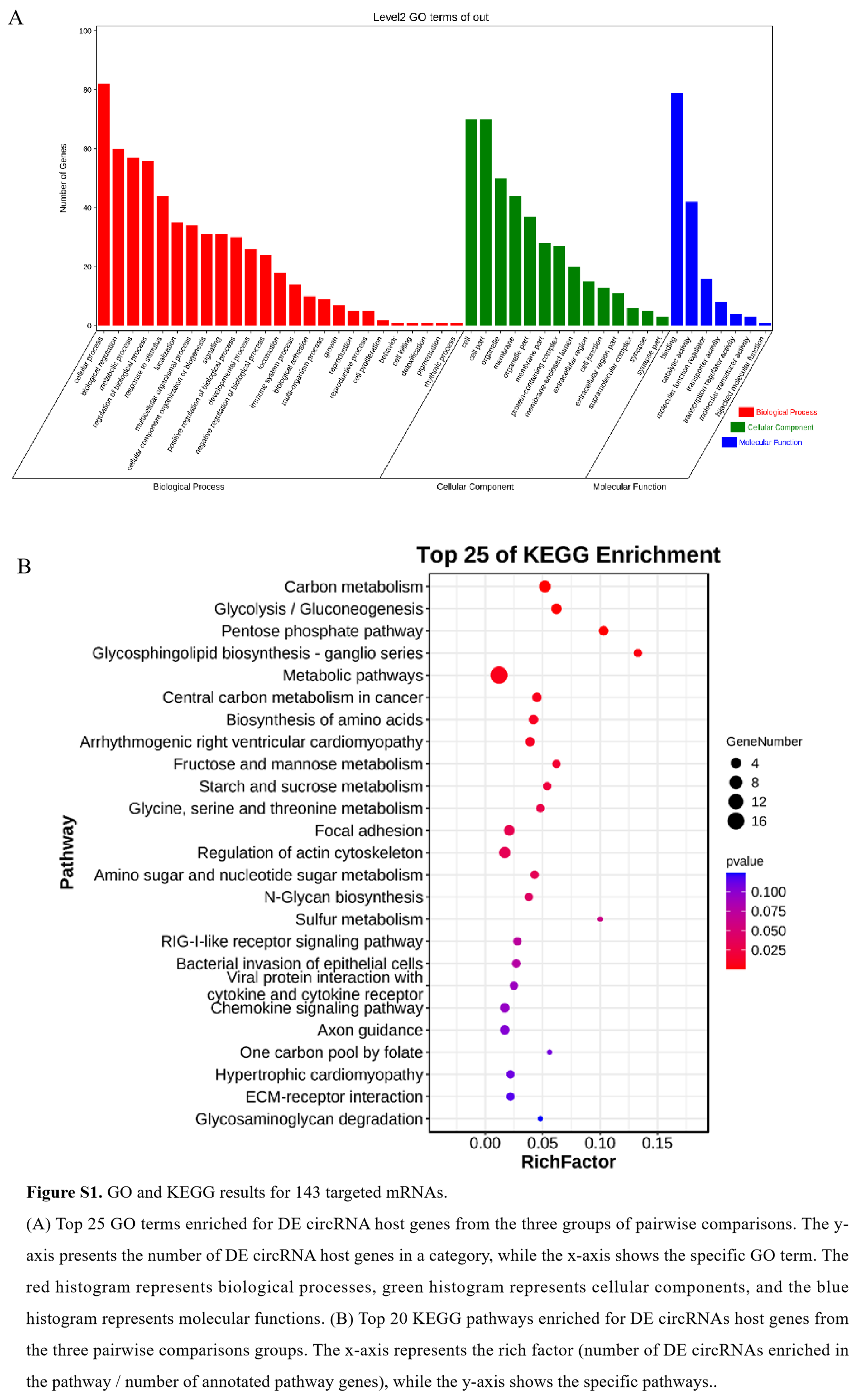
To explore the functional significance of mRNAs in the circRNA–miRNA–mRNA regulatory network, we performed functional enrichment analysis on 143 targeted mRNAs. As a result the GO enrichment analysis showed that these target mRNAs were significantly enriched in pyruvate metabolism, hexose metabolism, striated muscle contraction, glycolysis process, muscle system process, and muscle contraction. More specifically, PGAM2, ALDOA, ACTN3, PKM, and GPI are involved in the process of glycolysis, and ACTN3, PI16, WDR1 are involved in striated muscle development. It is worth noting that ACTN3 is also involved in the positive regulation of fast-twitch skeletal muscle fiber contraction, skeletal muscle fiber development, skeletal muscle tissue growth, and other important processes (Supplementary Figure S1A). Furthermore, KEGG enrichment analysis showed that the same target mRNAs were mainly enriched in carbon metabolism, glycolysis/gluconeogenesis, and regulation of the actin cytoskeleton. Specifically, PGAM2, ALDOA, GPI, and EFCAB7 are involved in glycolysis and gluconeogenesis, HOMER3 and PDPK1 are involved in the FOXO signaling pathway, and PDPK1, ITGA5, and ITGB4 are associated with the PI3K-AKT signaling pathway (Supplementary Figure S1B). These result suggested that these circRNAs in the ceRNA network many play important role in the muscle development.
Experimental Validation of circ_0018595
From the above network, we selected circ_0018595 for follow-up study, speculating that it might be involved in regulating growth and development of porcine skeletal muscle via the predicted circ_0018595/miR-1343/PGM1 axis (Figure 5A). Circ_0018595is an exonic circRNA generated from exons 3 and 4 of STT3B. It was amplified from longissimus dorsi cDNA using specific divergent primers, and the product was subjected to Sanger sequencing to validate the ligation site (Figure 6A). We also processed the total RNA extract with RNase R and performed qRT-PCR, revealing that circ_0018595 was more resistant to RNase R than STT3B mRNA and 18S rRNA as expected (Figure 6B). The circ_0018595 was expressed in various tissues, especially in lung, liver, skeletal muscle, and kidney (Figure 6C). We also examined the expression of circ_0018595 in muscle of Jinfen White pigs at 1, 90, and 180 days, and found it was significantly associated with developmental stage, with strong upregulation between 1 day and 90 days (P <0.05), and maintained at 180 days (Figure 6D). Furthermore, the results of a nucleocytoplasmic separation assay showed that circ_0018595 is primarily localized in the cytoplasm, which was consistent with its proposed role as a sponge for miRNAs (Figure 6E). Finally, the RNAhybrid website found that circ_0018595 and miR-1343 had potential binding sits, which was also found between miR-1343 and PGM1 mRNA transcript (Figure 6F). The function and mechanism experiment of circ_0018595in the skeletal muscle development will be carried out in the future work.
{kind=link}