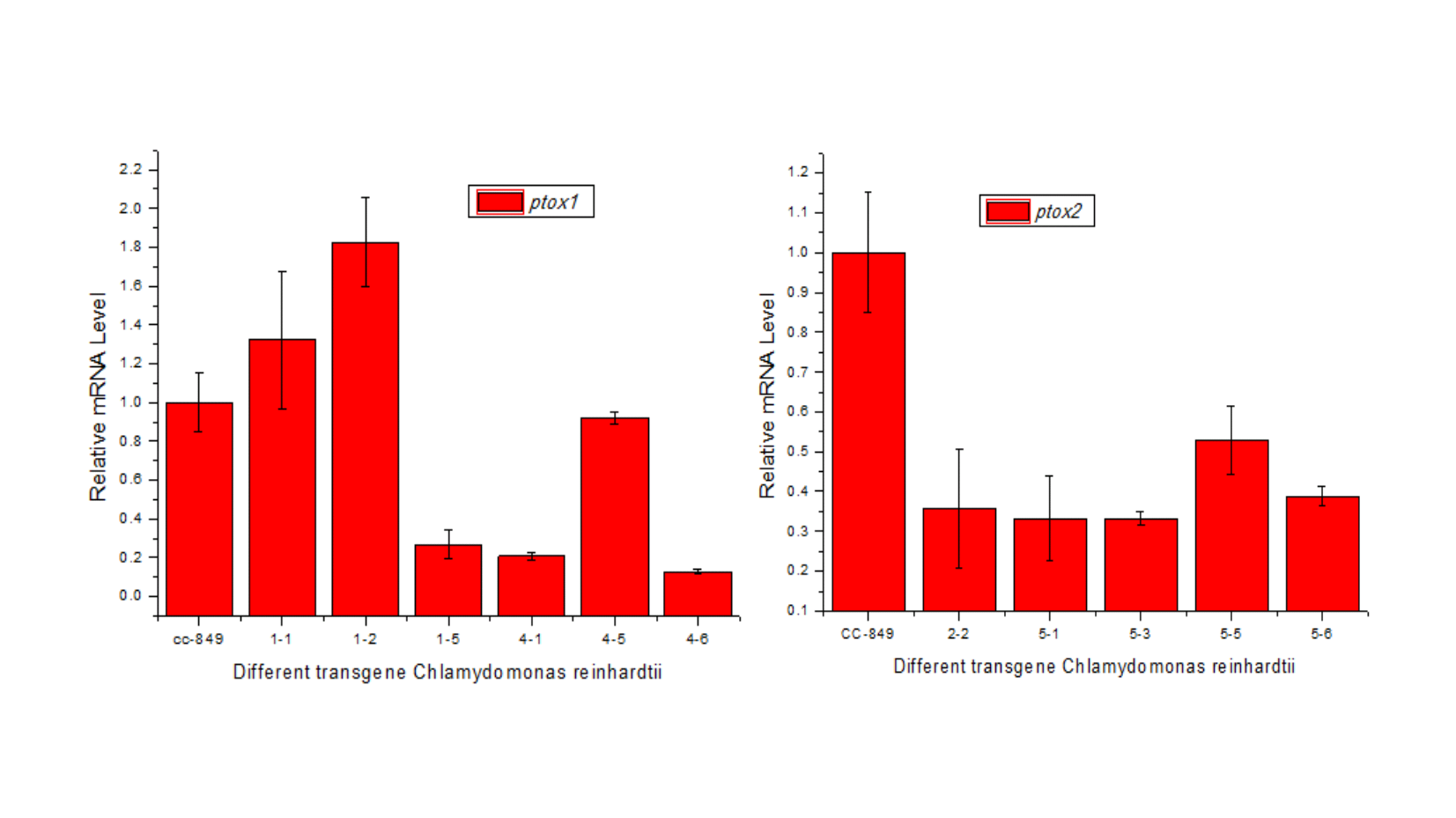
We established successfully two knock-down mutants, with 90% and 71% mRNA reduction of ptox1 and ptox2 under HL stress, respectively. Under normal culture condition, single knock-down of either PTOX did not inhibit cell growth, however, both mutants showed the significantly reduced growth under HL instead, more than 60% inhibition compared to only 30% in WT.
Current modification of PTOX expression, either knock-out or overexpression, shed lights on their multiple cellular functions. Knocking out PTOX in plants or microalgae resulted in severe phenotypes that encompass developmental and growth defects together with increased photosensitivity [36]. Interestingly, down-regulation of PTOX, approximately 3% of WT levels, did not compromise plant growth, under ambient growth conditions in Arabidopsis [37]. While over-expression of C. reinhardtii PTOX1 in plants makes the mutants more sensitive to HL than WT [38, 39] and Arabidopsis PTOX in tobacco promotes oxidative stress [40, 41]. Similarly, OsPTOX expression in Synechocystis did not affect growth under standard growth conditions (light intensities between 50 and 150 µmol photons m− 2 s− 1) [42]. In other stress treatments, over-expression of PTOX from the salt-tolerant brassica species Eutrema salsugineum show faster induction and a greater final level of PTOX activity once exposed to salt stress [43].
Based on mRNA level comparison between WT and a delta-psbA tobacco plant, up-regulation of the alternative electron transport pathways (NDH complex and PTOX) occurs at the translational or post-translational levels [44]. This suggests that PTOX is normally in excess, with delicate expression regulations in not only plants but also microalgae.
The differential expression patterns and the complementary relationship of ptoxs under both normal and HL treatments indicate complicate and different potential roles of these two PTOX genes in chlororespiration and stress responses. For instance, the astaxanthin accumulation level was much more in ptox2 silenced strain than ptox1, compared to WT. What’s more, significantly increased hydrogen production was observed in ptox1 silenced strain.
In Glycine max, differential expression of recently duplicated PTOX genes during plant development and stress conditions were extensively investigated [45]. The majority of plant species contain only a single gene encodes PTOX. Previously, two putative PTOX (PTOX1 and 2) genes were identified in Glycine max. In development, PTOX1 was predominant in young tissues, while PTOX2 was more expressed in aged tissues. Under stress conditions, the PTOX transcripts varied according to stress severity, i.e., PTOX1 mRNA was prevalent under mild or moderate stresses while PTOX2 was predominant in drastic stresses. Overall, the results indicate a functional relevance of this gene duplication in G. max metabolism, whereas PTOX1 could be associated with chloroplast effectiveness and PTOX2 to senescence and/or apoptosis [45].
The differential expression of ptoxs was also observed in Chlamydomonas under phosphate deprivation where ptox2 mRNA level was up-regulated about 13-fold whereas the ptox1 transcripts increased 2.4-fold after 48 h of treatment [32]. In a knockout mutant of PTOX2 in Chlamydomonas the plastoquinone pool is constitutively reduced under dark-aerobic conditions, and the ptox2 mutant shows lower fitness than wild type when grown under phototrophic conditions [33]. In another report, the absence PTOX2 and cytochrome b6f complex of C. reinhardtii, results in almost complete reduction of the plastoquinone pool in light [34]. In this study, both ptox genes connected with cell growth inhibition, but with no significantly difference. Interestingly, under normal light intensity, ptox1 mRNA levels correlated with the growth in both WT and PTOX1i mutant while ptox2 remained relatively low under both WT and PTOX2i strains in this study. Thus, ptox1 may involve in cell growth and ptox2 plays other metabolic roles in Chlamydomonas.
PTOX is regarded as an enzyme at the crossroads of various metabolic processes, such as regulation of cyclic electron transfer and carotenoid biosynthesis [36]. PTOX is very important for carotenoid biosynthesis, since the phytoene desaturation, a key step in the carotenoid biosynthesis, is blocked in the white sectors of Arabidopsis im mutant [3]. The absence of PTOX in plants usually results in photo-bleached variegated Arabidopsis leaves [46] and impaired adaptation to environment alteration, and mutant plants will not survive the mediocre light intensity during its early development stage [3]. Although PTOX level and activity has been found to increase under a wide range of stress conditions [9].
Thus, PTOX involves in carotenoid biosynthesis but which ptox gene plays more important role in this process in microalgae is still under investigation.
In other green microalga Haematococcus, Hptox1 and Hptox2 also showed differential expression patterns in response to various oxidative stresses [20]. And the authors regarded Hptox1 as the key PTOX gene for co-regulation of astaxanthin accumulation in Haematococcus [20]. The rice ptox1 mutant accumulated phytoene in white leaf sectors with a corresponding deficiency in beta-carotene, consistent with the expected function of PTOX1 in promoting phytoene desaturase activity. Our results demonstrate that PTOX1 is required for carotenoid synthesis [47].
Similar to heat and drought, HL treatment stimulates chlororespiration in higher plants [48] and microalgae [20], causing the up-regulation of the PTOX and the thylakoidal NADH DH complex [49, 50]. The natural astaxanthin mainly derives from the microalgae producer, Haematococcus. The induction of nitrogen starvation and high light intensity is particularly significant for boosting astaxanthin production [51].
Under HL, ptox1 mRNA level was up-regulated significantly (more than 10 fold compared to WT) in PTOX2i mutant. Together with our study, it suggests that ptox expressions differ temporally or spatially in response to various stressors. We speculated that it is ptox1 rather than ptox2 that is co-regulated, or functionally coupled with carotenoid biosynthesis in Chlamydomonas, similar to Haematococcus.
Hydrogen production in green mciroalgae requires electrons from the photosynthetic electron transfer chain to reduce H+. Sulfur-deprived cultivation of C. reinhardtii [52] was previously regarded as the most efficient technique to enhance photobio-H2 production in microalgae [53–55]. Most recently, modification of photosynthetic genes and even non-coding RNAs in Chlamydomonas significantly improve the bio-hydrog8 en production [35, 56, 57].
Under HL, ptox2 responds to the stress with delayed maximum expression level in ptox1 silenced strain-PTOX1i mutant, with a similar but delayed up-regulation level with WT. What’s more, significantly increased hydrogen production was observed in the PTOX1i mutant. We address the importance of ptox2 gene in hydrogen bioproduction in Chlamydomonas.
The limitations of our study are the absence of evidence based on protein levels, and RNAi knock-down still remains relatively high basal expression of the target genes. Further investigations involving in Western blotting and genome editing (for complete individual or double knock-out, if possible) would be highly required for a better understanding of the diverse functions of PTOX genes in microalgae.
{kind=link}
{kind=link}
{kind=link}