HFD Feeding Induces Obesity and Glucose Intolerance in Our DIO Model
To test whether HFD feeding also decreases sorcin expression in the hypothalamus of mice as in their pancreatic islets of Langerhans (Marmugi et al), we used male C57BL/6J mice fed either a control normal chow (NC) diet or a HFD from Research Diets™, containing 60% kcal from fat (D12492).
Mice were placed on a NC or HFD at 5 weeks of age and body weight measurements taken regularly to confirm development of obesity in HFD fed mice. In addition to this, body composition was assessed using EchoMRI at the end of the dietary intervention. IPGTTs were conducted at 8, 10 and 16 weeks old to confirm development of glucose intolerance and metabolic dysfunction.
As shown in Fig. 1A, mice fed a HFD gained weight significantly more quickly than mice on SD, with significant weight differences developing 5 weeks post HFD introduction (Fig. 1A: 10 weeks of age, SD = 25.78 ± 0.37g vs HFD = 31.25 ± 0.93g, n = 6, p < 0.05, 2-way ANOVA, Šídák's multiple comparisons test). These differences in weight gain increased with time and at the end of the 13-week study, HFD fed mice on average weighed 12.70g more than SD fed mice (Fig. 1A: 18 weeks of age, SD = 31.13 ± 0.48g vs HFD = 43.83 ± 2.42g, n = 6, p < 0.0001, 2-way ANOVA, Šídák's multiple comparisons test). Total body fat mass was also measured 13 weeks after diet administration, showing significant differences in body-fat mass between SD and HFD fed mice, suggesting that increased body fat accounts for most of the difference in weight between the two groups. HFD fed mice show 20.03% increase in body-fat percentage compared with SD fed mice, with a body fat percentage of 40.00 ± 2.75% (Fig. 1B: 18 weeks of age, SD = 19.97 ± 0.57% vs HFD = 40.00 ± 2.75%, n = 6, p < 0.0001, analysis with unpaired two-tailed t-test).
As shown in Fig. 2, intraperitoneal glucose tolerance tests (IPGTTs) using 1g glucose/kg body weight were conducted 3 weeks, 5 weeks and 11 weeks post dietary intervention and show a gradual but significant worsening of glucose tolerance throughout this period. HFD fed mice exhibit slightly but significantly altered glucose tolerance at 8 weeks of age, only 3 weeks after dietary intervention when challenged with 1g/kg glucose (Fig. 3A: SD AUC = 1368 ± 52.99 AU vs HFD = 1577 ± 65.44 AU, n = 6, p < 0.05, AUC analysis with unpaired two-tailed t-test). 5 weeks after dietary intervention, HFD fed mice exhibited markedly worsened glucose tolerance, which was exacerbated at 11 weeks post dietary intervention (Fig. 3B: 5 weeks after intervention, SD AUC = 1465 ± 75.71 AU vs HFD AUC = 1800 ± 57.32 AU, n = 6, p < 0.01; Fig. 3C: 11 weeks after intervention, SD AUC = 1377 ± 45.00 AU vs HFD AUC = 2079 ± 168 AU, n = 6, p < 0.01, AUC analysis with unpaired two-tailed t-test). Work by the Jackson Laboratory with C57BL/6 mice found comparable body fat percentage and weight at 16 weeks of age, associated with increased triglyceride concentrations, hyperinsulinemia and hyperleptinemia phenocopying early stage Type 2 Diabetes and metabolic syndrome (The Jackson Laboratory, 2022).
Sorcin Expression Remains Unchanged in The Arcuate Nucleus of HFD fed Mice
As HFD feeding has been previously linked with hypothalamic ER stress and its association with leptin resistance (Ozcan et al.) we wanted to determine whether reduced expression of sorcin might be responsible for these effects. To determine this, after 13 weeks of dietary intervention, our DIO and control mice were culled and brains were then dissected to isolate the arcuate nucleus (ARC) and RNA extracted for downstream qPCR analysis of gene expression. The results reveal non-significant differences in mRNA levels of sorcin in the ARC (Fig. 3A sorcin: SD = 1.00 ± 0.07 vs HFD = 0.99 ± 0.06, n = 5 vs n = 4, p = 0.98, analysis with unpaired two-tailed t-test). Also, there was no changes in the expression of ER stress marker CHOP and XBP1s in the ARC (Fig. 3B CHOP: SD = 1.00 ± 0.13 vs HFD = 1.09 ± 0.05, n = 5 vs n = 4, p = 0.60, analysis with unpaired two-tailed t-test, Fig. 3C XBP1s: SD = 1.00 ± 0.16 vs HFD = 1.34 ± 0.14, n = 5 vs n = 4, p = 0.17, analysis with unpaired two-tailed t-test).
C57BL/6 Sorcin Null Mice Show Unchanged Body Weight and Fat Mass on HFHSD
Wild-type (Sri+/+), heterozygous (Sri+/−) and sorcin null mice (Sri−/−) on C57BL/6J genetic background were placed on a HFHSD at 6 weeks of age and body weight measurements taken weekly to determine whether sorcin null mice showed increased sensitivity to dietary challenge. We found non-significant differences in body weight between across the 12 weeks of HFHSD feeding (Fig. 4A: 18 weeks of age: Sri+/+ = 38.91 ± 2.29g vs Sri+/− = 38.66 ± 1.49g vs Sri−/− = 38.92 ± 1.38g, n = 10 vs 16 vs 12, Sri+/+ vs Sri+/− p = 0.99, Sri+/+ vs Sri−/− p = 0.99, Sri+/− vs Sri−/− p = 0.99, 2-way ANOVA, Šídák's multiple comparisons test).
We also measured body fat percentage at 18 weeks of age using EchoMRI (Fig. 4B). These results show a tendency towards increased body fat mass in heterozygous and sorcin null animals, but results are heavily negatively skewed in wild-type mice by three animals and thus show significant deviation (Sri+/+ = 31.81 ± 4.32% vs Sri+/− = 38.51 ± 1.41g vs Sri−/− = 39.36 ± 1.27g, n = 7 vs 14 vs 10, Sri+/+ vs Sri+/− p = 0.10, Sri+/+ vs Sri−/− p = 0.08, Sri+/− vs Sri−/− p = 0.95, 2-way ANOVA, Šídák's multiple comparisons test).
C57BL/6 Sorcin Null Mice Exhibit Non-Significant Changes in ARC ER Stress Marker Expression
To determine whether sorcin knockout lead to an increase in ER stress in the ARC nucleus, the main effector of leptin signalling, we measured expression of ER stress markers in RNA isolated from the ARC nucleus of these animals.
As expected, sorcin relative expression was significantly lower in the heterozygous (Sri+/−) and sorcin null mice (Sri−/−) compared to wild-type (Sri+/+) mice (Sri+/+ = 1.00 ± 0.01 vs Sri+/− = 0.66 ± 0.02 vs Sri−/− = 0.17 ± 0.02, n = 5 vs 12 vs 10, Sri+/+ vs Sri+/− p < 0.0001, Sri+/+ vs Sri−/− p < 0.0001, Sri+/− vs Sri−/− p < 0.0001, 1-way ANOVA, Tukey's multiple comparisons test). However, we found non-significant changes in the ER stress markers CHOP, ATF6 and XBP1s in Sri+/− and Sri−/− mice compared to wild-type littermate controls. There was however a tendency towards increased expression of XBP1s suggesting mildly increased ER stress which demands further investigation (Sri+/+ = 1.00 ± 0.02 vs Sri+/− = 1.31 ± 0.11 vs Sri−/− 1.34 ± 0.12, n = 5 vs 11 vs 9, Sri+/+ vs Sri+/− p = 0.01, Sri+/+ vs Sri−/− p = 0.03, Sri+/− vs Sri−/− p = 0.96, 1-way ANOVA, Tukey's multiple comparisons test).
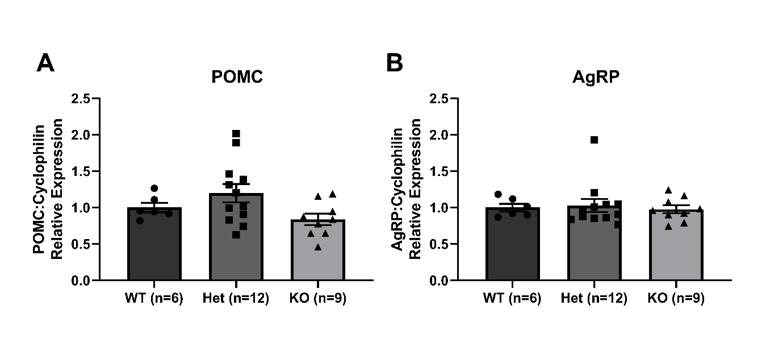
We also investigated changes in expression of the orexigenic and anorexigenic neuropeptide markers AgRP and POMC respectively, to determine if this highlighted any changes in leptin signalling. Our results show non-significant changes in expression of either neuropeptide, consistent with the unchanged body weight and food-intake (Suppl. Figure 1 POMC: Sri+/+ = 1.00 ± 0.06 vs Sri+/− = 1.20 ± 0.13 vs Sri−/− 0.84 ± 0.08, n = 6 vs 12 vs 9, Sri+/+ vs Sri+/− p = 0.47, Sri+/+ vs Sri−/− p = 0.63, Sri+/− vs Sri−/− p = 0.06; AgRP: Sri+/+ = 1.00 ± 0.06 vs Sri+/− = 1.20 ± 0.13 vs Sri−/− 0.84 ± 0.08, n = 6 vs 12 vs 9, Sri+/+ vs Sri+/− p = 0.47, Sri+/+ vs Sri−/− p = 0.63, Sri+/− vs Sri−/− p = 0.06, 1-way ANOVA, Tukey's multiple comparisons test)
C57BL/6 Sorcin Null Mice Show a small increase in food Intake after leptin administration
Given the role of hypothalamic ER stress in leptin resistance, we also wanted to determine changes in leptin sensitivity in Sri−/− mice. We attempted to do this using pSTAT3 staining of perfusion-fixed coronal brain slices after leptin administration but were unable to optimise the protocol in our mice. Therefore, as an indirect measure of leptin sensitivity, we measured both ad-libitum food intake and food intake after leptin administration in our cohort of mice, on standard chow diet in 8 w/o mice (Fig. 6A, B) or after feeding a HFHSD in 20 w/o mice for 12 weeks (Fig. 6C, D). We observed non-significant differences at all time-points between Sri+/+, Sri+/− and Sri−/− mice in ad-libitum food intake at both 8 and 20 weeks of age. Food intake after leptin administration (3 µg/g) also showed no significant difference at most time-points analysed; cumulative food intake at 8 hours after leptin administration was slightly increased in Sri−/− mice at 8 weeks of age (Fig. 6B: Sri+/+ = 2.37 ± 0.35g vs Sri+/− = 2.90 ± 0.29g vs Sri−/− = 3.08 ± 0.28g, n = 3 vs 9 vs 6, Sri+/+ vs Sri+/− p = 0.10, Sri+/+ vs Sri−/− p = 0.03, Sri+/− vs Sri−/− p = 0.65, 2-way ANOVA, Šídák's multiple comparisons test), but this observation disappeared at the 24-hour time point.
Sorcin Null HEK293-LepRb Cells Show Unchanged Leptin Sensitivity
As previously mentioned, other studies have measured the effects of sorcin depletion or overexpression on STAT3 signalling using in-vitro models. Li et al. found that sorcin overexpression in the hepatocellular carcinoma cell line HepG2 increases STAT3 phosphorylation in response to IL-6 stimulation (Li et al., 2017). To determine whether similar mechanisms might occur in response to leptin stimulation, we used our HEK293 sorcin null cells expressing the long-isoform of the leptin receptor (LepRb) by transient transfection with LepRb-HA plasmid. These cells were then stimulated with leptin (0–10 ng/mL), and STAT3 phosphorylation (Tyr705) measured by western blotting of cell lysates. HEK293 wild-type cells and non-transfected cells were used to control against sorcin and LepRb independent STAT3 signalling.
As shown in Fig. 7, there was no significant difference in pSTAT3/STAT3 ratio between wild-type and sorcin null HEK293 cells for any concentration of leptin stimulation. Rather than reduced leptin signalling in sorcin null cells, there is a tendency for increased leptin signalling, with sorcin null cells showing consistently higher pSTAT3/STAT3 ratios for all concentrations of leptin stimulation. Despite this, two-way ANOVA analysis shows non-significant differences between genotypes analysed across all data (wild-type vs Sri null p = 0.15).
{kind=link}