In search for potential fire blight control agents, we isolated E. amylovora bacteriophages VyarbaL and Hena2 from natural sources on the territory of Belarus. The VyarbaL phage was isolated from the soil sample collected under a tree of the genus Salix (Belarus, Rudensk, 2018). Phage Hena2 was obtained from the soil under apple and pear trees collected in a private courtyard (Belarus, Novogrudok, 2017). Bacteriophages VyarbaL and Hena2 form clearly distinguishable plaques when plated using the double-layer agar technique. VyarbaL forms plaques of 3–3.5 mm in diameter with a well-marked halo; Hena2 plaques were smaller, from 0.5 mm to 2 mm in diameter, with jagged edges. When the drops of the concentrated (107 PFU mL− 1) phage suspension of VyarbaL or Hena2 were applied onto the lawn of E. amylovora 1/79Sm, turbid lysis zones were observed.
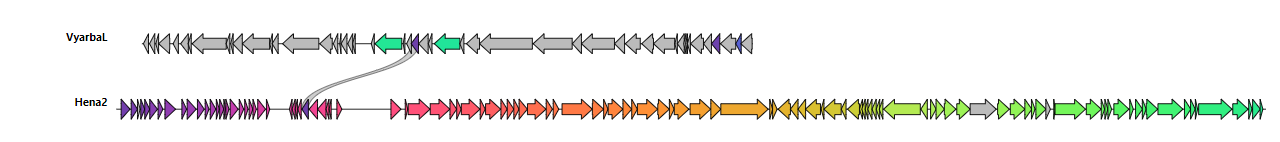
Genomic DNA was extracted from DNAse treated lysates of bacteriophages VyarbaL and Hena2, and complete genome sequences were determined. Double-stranded DNA genomes of bacteriophages had different sizes: 44768 b.p. (VyarbaL) and 84251 b.p. (Hena2). Using PhageTerm analysis, the headful mode of packaging PAC was concluded for VyarbaL with a defined terminus only on one strand and approximative redundancy of 298 bp. The Hena2 genome has direct terminal repeats (439 bp) and obvious termini.
In the VyarbaL genome 49 open reading frames (ORF) were predicted; no tRNA genes were found (Fig. 1). The genome of the bacteriophage Hena2 encoded 114 ORFs and 26 tRNAs genes. All the genes of the VyarbaL genome have the same orientation whereas Hena2 genes organized in several clusters with different orientations. GC-content of the VyarbaL genome 49.8%, Hena2–43.5% (Fig. 1).
Bacteriophage genome sequences were queried against the viruses (taxid:10239) nucleotide collection (nr/nt) using blastn (carried out on 06/05/2022). Sequence similarity searches revealed that VyarbaL presents 98.44% (coverage 98%) sequence identity with Erwinia phages vB_EamP-S2 (NC_047917.1). The bacteriophage Hena2 genome had 99.42% (coverage 100%) of similarity to the phage vB_EamM-M7 (NC_041978.1). Thus VyarbaL (NCBI:txid2923252) is a podovirus that can be classified in the Caudoviricetes class, the Autographiviridae family, within the Molineuxvirinae subfamily, shares the Eracentumvirus S2 species with the phage vB_EamP-S2. Bacteriophage Hena2 (NCBI:txid2923253) is a myovirus, classified in the Caudoviricetes class, within the Ounavirinae subfamily, sharing the Kolesnikvirus M7 species with vB_EamM-M7 virus.
Global sequence alignment of the sequences of the proteins encoded by these two bacteriophages using clinker (identity threshold 0.3) showed that VyarbaL and Hena2 share only one similar protein - lysin, sharing 35.03% amino acid identity (82% coverage) (Supplementary material). The two bacteriophages have genes with the same function: predicted structural genes coding the major capsid protein and tail fiber protein; - holin and endolysin genes, involved in host lysis by bacteriophages; and the packaging gene synthesizing terminase. The bacteriophages possess DNA polymerase, exonuclease, endonuclease and DNA ligase genes, involved in the replication and metabolism of DNA and RNA. The genomes differ in the presence of genes for specific structural and assembly proteins, that reflect different types of the virion organization. The bacteriophage Hena2 genome also contains a diverse set of putative transferases and oxidoreductases. In both bacteriophages we were able to identify genes responsible for the specific interaction between the phage and the host. The VyarbaL bacteriophage gene encodes the EPS-depolymerase enzyme, which explains the presence of a distinct halo around bacteriophage plaques. Note that known E. amylovora bacteriophages differ in the Efficiency of plating on strains with different levels of EPS production [32]. The presence of this enzyme in VyarbaL suggests the possibility of overcoming the barrier of the extracellular matrix for interaction with the cell surface.
The RIIA and RIIB genes in the Hena2 genome are reported to confer to the T4 phage the ability to escape Rex exclusion by a λ lysogen [33].
Given the absence of any identifiable genes associated with the lysogenic life cycle in VyarbaL and Hena2 genomes, the inherence of turbid plaques indicates that some mechanisms allow significat fraction of the potentially sensitive host cell to survive the exposure to high bacteriophage concentration and to give rise to the microcolonies. This effect may allow the phage to form long lasting metastable associations with the host in which a fraction of the cells gets lyzed to maintain the bacteriophage reproduction while the remaining fraction of bacteria is somehow (temporally) protected from the phage attack to enable bacterial growth. To test this hypothesis, we analyzed the ability of these two bacteriophages to form the PA.
The PA of the VyarbaL bacteriophage and the E. amylovora 1/79Sm bacterial culture were shown to be stable over multiple passages. After 14th passage the bacteriophage active against the parental host strain was present in 90% of the PAs (Fig. 2, supplementary material). Bacteriophage DNA was detected by PCR analysis for the main capsid protein gene (gMCP) of the phage VyarbaL in 95% of the PAs after 14th passage (Fig. 3).
In contrast to VyarbaL system, all but one (#11) PAs formed with the Hena2 bacteriophage lost the phage by the eights passage. The data are presented in Fig. 4 and supplementary material. However the PA #11 produced the phage till the fourteenth passage and was PCR-positive for the gMCP of Hena2 (Fig. 5).
Simultaneously with performing the passages, we determined the CFU, PFU and free PFU counts at the passages 5, 8, 11 and 14 for three randomly chosen PA from each phage-host system (these were PA ## 2, 14 and 16 for VyarbaL system and ##10, 11 and 20 for Hena2 system). The ratio of the total PFU to CFU counts in three randomly selected PAs of VyarbaL system for a series of passages varied from 10− 2 to 6 x 10− 4. The ratio of PFU of free bacteriophage to CFU for the PA of the VyarbaL system in a number of passages ranged from 10− 2 to 10− 4 (Fig. 6).
For the PA of the Hena2 system, the PFU / CFU ratio was higher compared to the VyarbaL system, ranging from 10− 1 to 100. However, the PAs # 10 and #20 lost the bacteriophage after the 8th and 7th passages, respectively.
The PA are likely to represent co-evolving systems in which both host and phage genotypes and/or phenotypes may differ from the parental bacterial and viral strains [34, 35]. To determine the features of the components we re-isolated the evolved phages from the last (14th) passage from the PAs ## 14 and 16 of the VyarbaL system and from the only PA (#11) of the Hena2 system retaining the phage by the 14th passage. We also obtained 32 bacterial subclones from each of two VyarbaL PAs and 64 subclones from the Hena2 PA #11. These subclones were spot-tested for the sensitivity towards the corresponding original phages and the evolved phages derived from the same PA. In all the cases the bacteria appeared resistant, although some of the Hena2 PA sublcones showed very turbid inhibition zones with both original and evolved phages. However, when phage dilutions were plated no plaques were visible.
We then cross-tested a subset (n = 14) of the subclones from the PAs with VyarbaL and Hena2 phages for the sensitivity to the non-parental phages (Hena2 and VyarbaL respectively). In all the cases the subclones remain sensitive indicating that the mechanisms of the resistance to these two viruses were different (Fig. 8).
It is known that LPSs have great structural diversity and determine many serotypes of Gram-negative bacteria. Also, in some cases, LPS serve as receptor molecules for bacteriophages [36, 37]. To determine if the O antigen alterations are involved in the observed resistance we determined the LPS profiles of the parental strains and 7 subclones from each of the systems studied (Fig. 9) but no differences were observed.
{kind=link}