Endogenous AGO1 immunoprecipitation for the in vitro RISC activity assay
By using endogenous AGO1, the in vitro RISC assay can eliminate the limitation of plant material; however, a good quality a-AGO1 antibody, which did not affect AGO1 function, would determine whether the in vitro RSIC assay can be successfully performed. Although commercial AGO1 is available, we made a homemade a-AGO1 antibody because we needed large amounts of AGO1 to set up the assay. Moreover, the generation know-how can be applied to producing other AGO antibodies, e.g., AGO2.
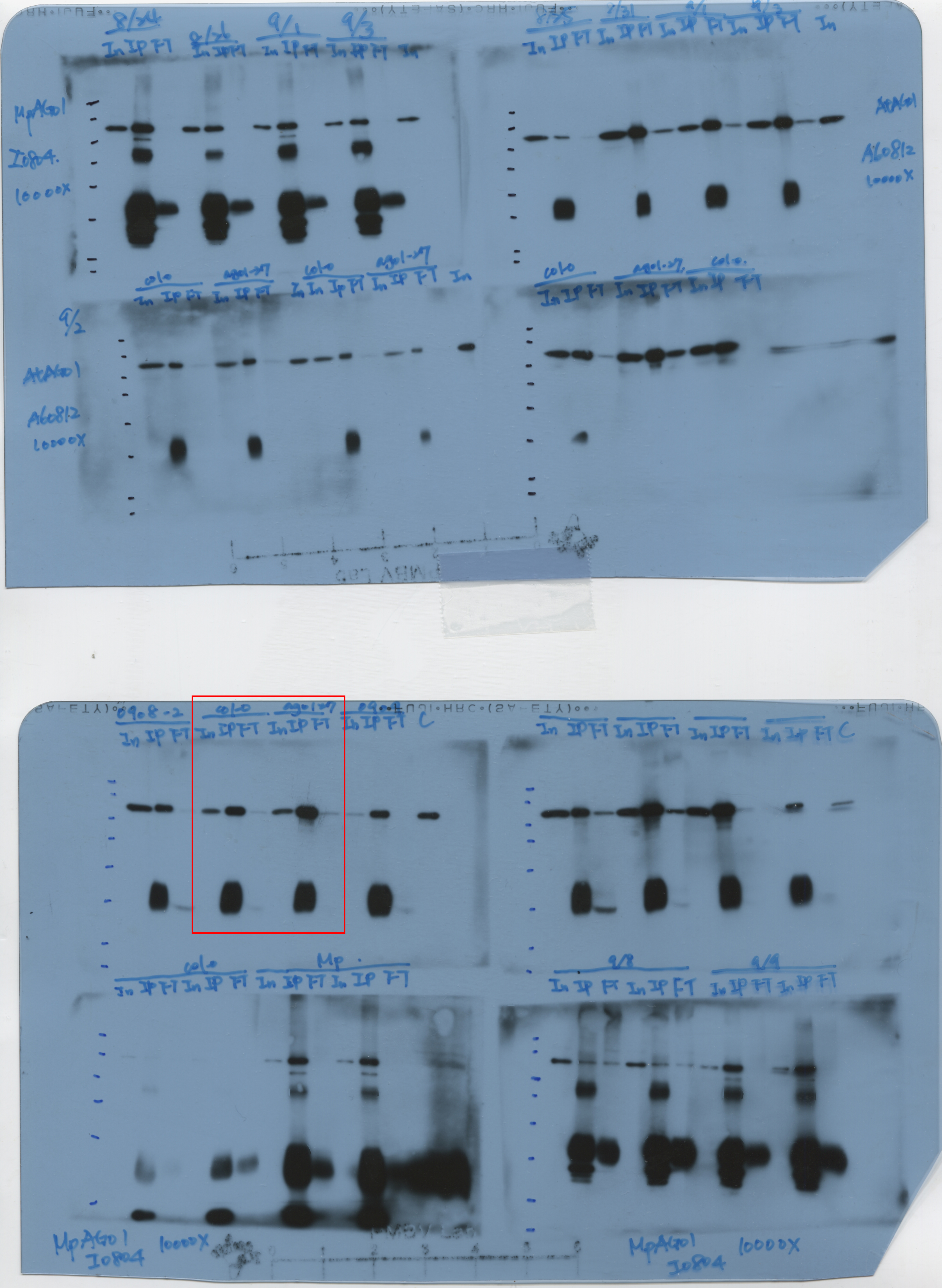
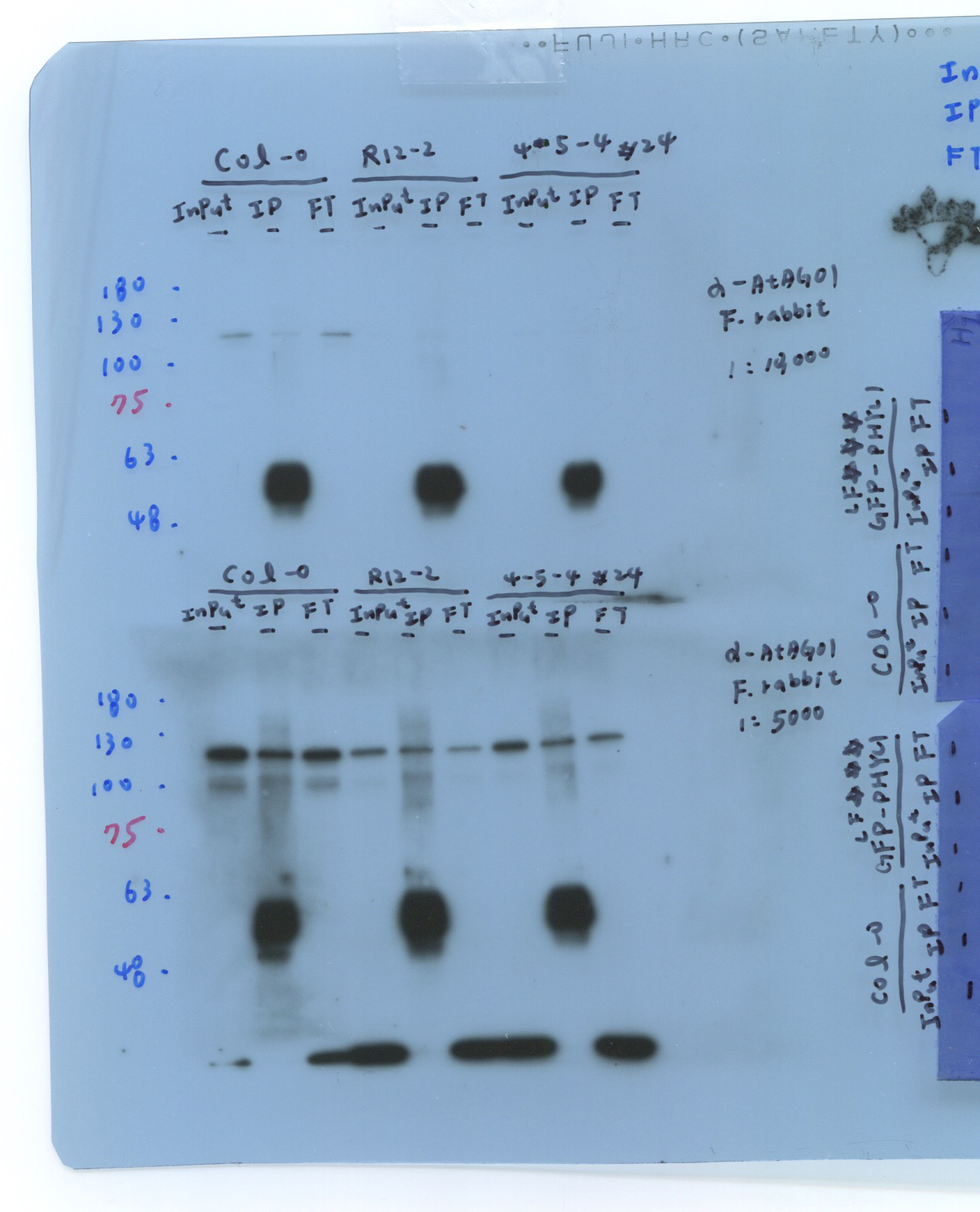
The amino acid alignments of AGO1, AGO2, AGO4, and AGO10 showed that the 1-256 aa region is variable, with 12.2%-30.5% identity (Fig. 1a). The N-terminus of AGO1 (1-256 aa) was more similar to the N-terminus of AGO10 (1-205 aa), with a shared identity of 30.5%. Thus, the N-terminus of AGO1 (1-240 aa) was selected for recombinant protein production (his-AGO1N240) and purified as an antigen to the AGO1 antibody (Fig. 1a, box). Western blot analysis showed that the a-AGO1 antibody could recognize endogenous AGO1 (117 kDa) in Col-0 plants. In contrast, the AGO1 signal was absent in the ago1-36 null mutant, suggesting that the homemade a-AGO1 antibody can be tested for its IP ability in an in vitro RISC assay (Fig. 1b).
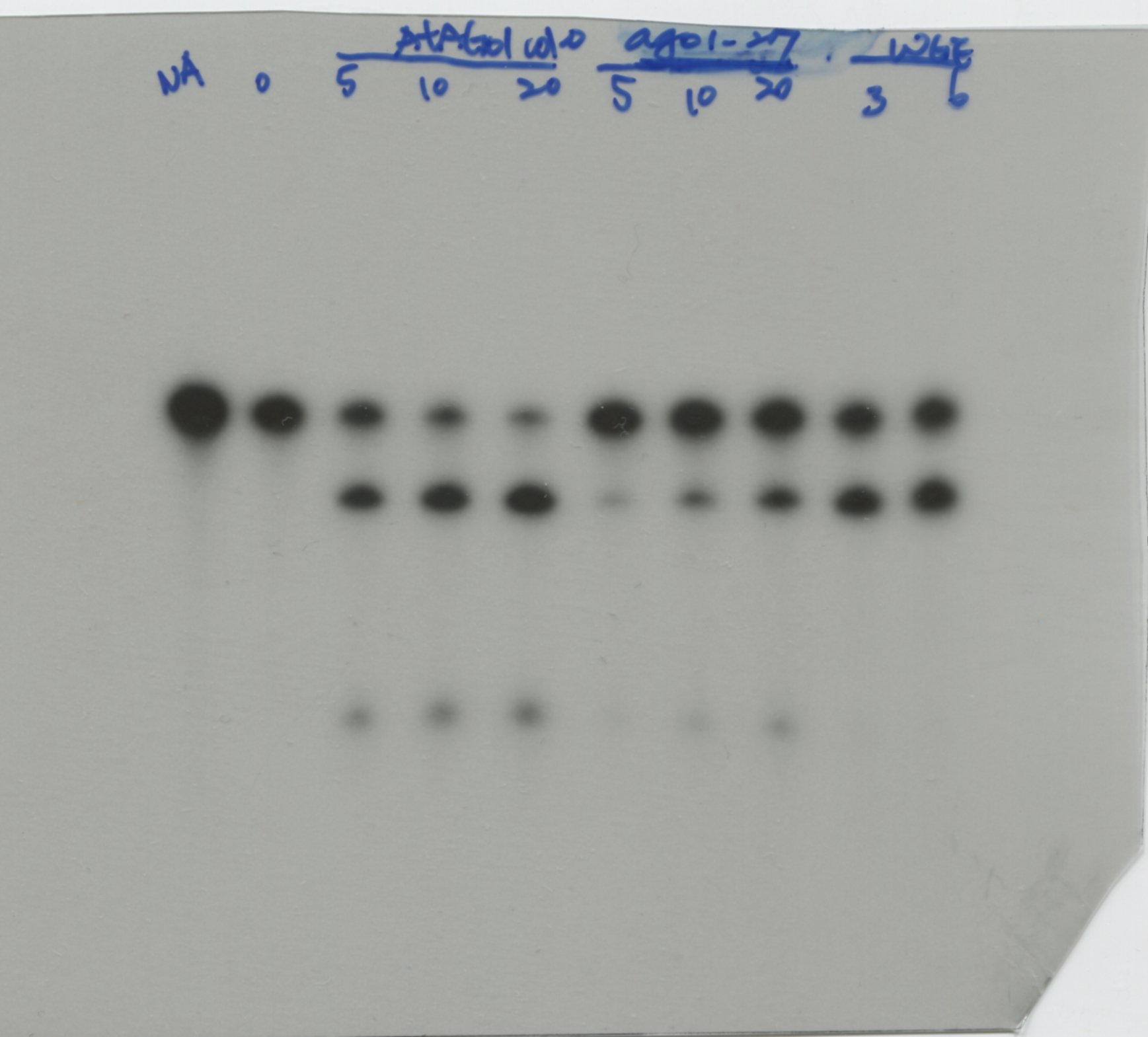
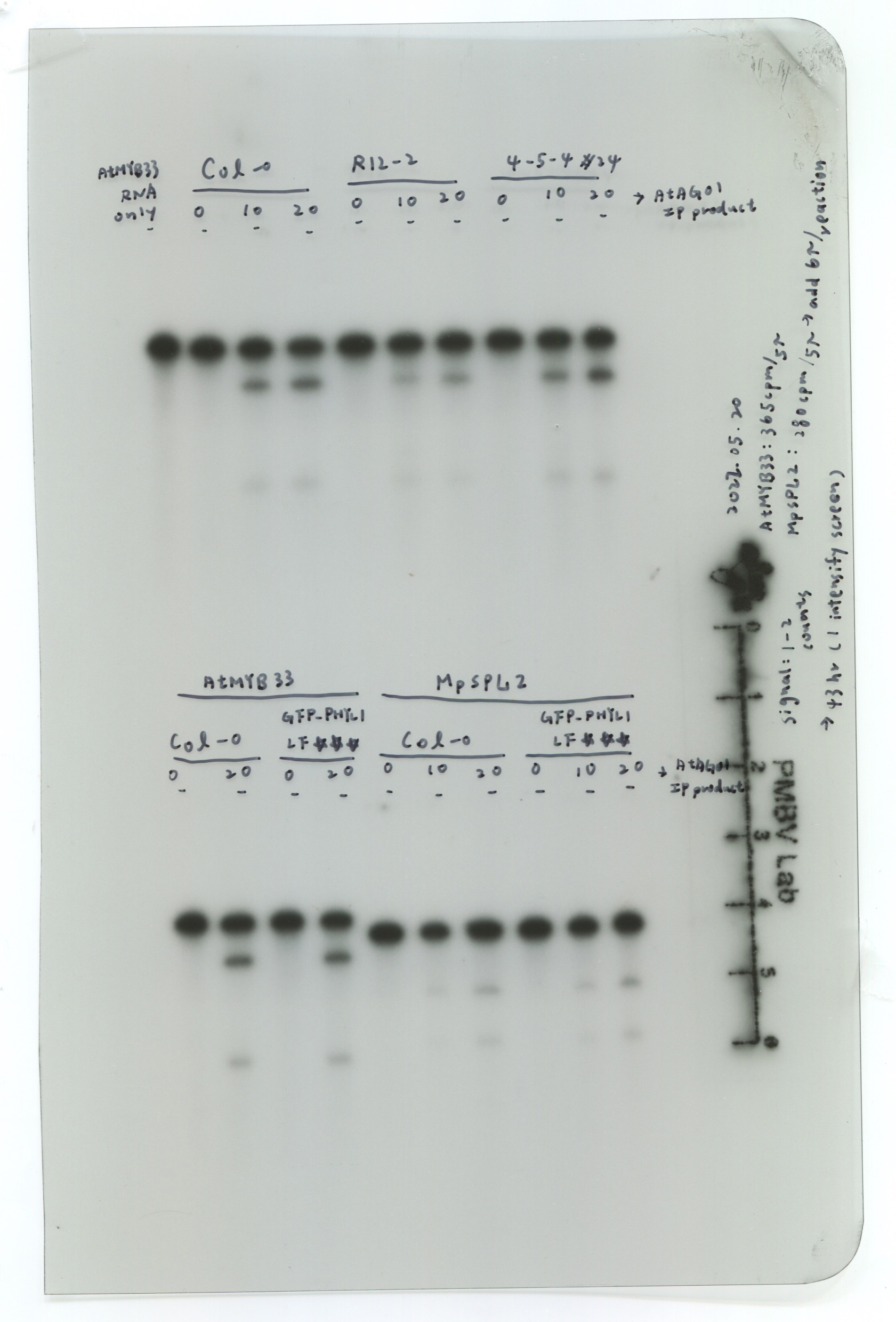
To evaluate the in vitro RISC assay with the a-AGO1 antibody, endogenous AGO1 was immunoprecipitated from Col-0 seedlings and ago1-27 mutants and assessed with the RNA substrate MYB33 (MYB33-230), which contained a target site for miR159a and miR159b (Fig. 2a). MYB33-230 is a substrate RNA (230 nt in length) that was generated by in vitro transcription (Fig. 2a). The AGO1-IP from Col-0 seedlings can target and cleave MYB33-230 (Fig. 2b). The signals of 5'- and 3'-cleaved fragments were increased when the amounts of AGO1-IP were increased, suggesting that AGO1 cleavage is dose-dependent (Fig. 2b). The ago1-27 mutant, with an Ala992Val weak allele produced by ethyl methane sulfonate mutation, showed attenuated slicer activity (Morel et al., 2002). AGO1-IP from the ago1-27 mutant revealed inefficient MYB33-230 cleavage (Fig. 2b). The amounts of AGO1-IP from Col-0 plants and ago1-27 mutants were compatible (Fig. 2c). The 20 ng of AGO1-IP from Col-0 plants resulted in 81% cleavage efficiency. In contrast, only 37% cleavage efficiency was observed in 20 ng AGO1-IP of ago1-27 mutants (Fig. 2d). We further mutated the target site in the seed region (MYB33mSeed) and center region (MYB33mCenter) (Fig. 2e). The results showed no RISC cleavage of MYB33mSeed or MYB33mCenter RNA substrates, whereas MYB33-230 was cleaved as a positive control (Fig. 2f). These results indicated that the a-AGO1 antibody could be used for the in vitro RISC assay.
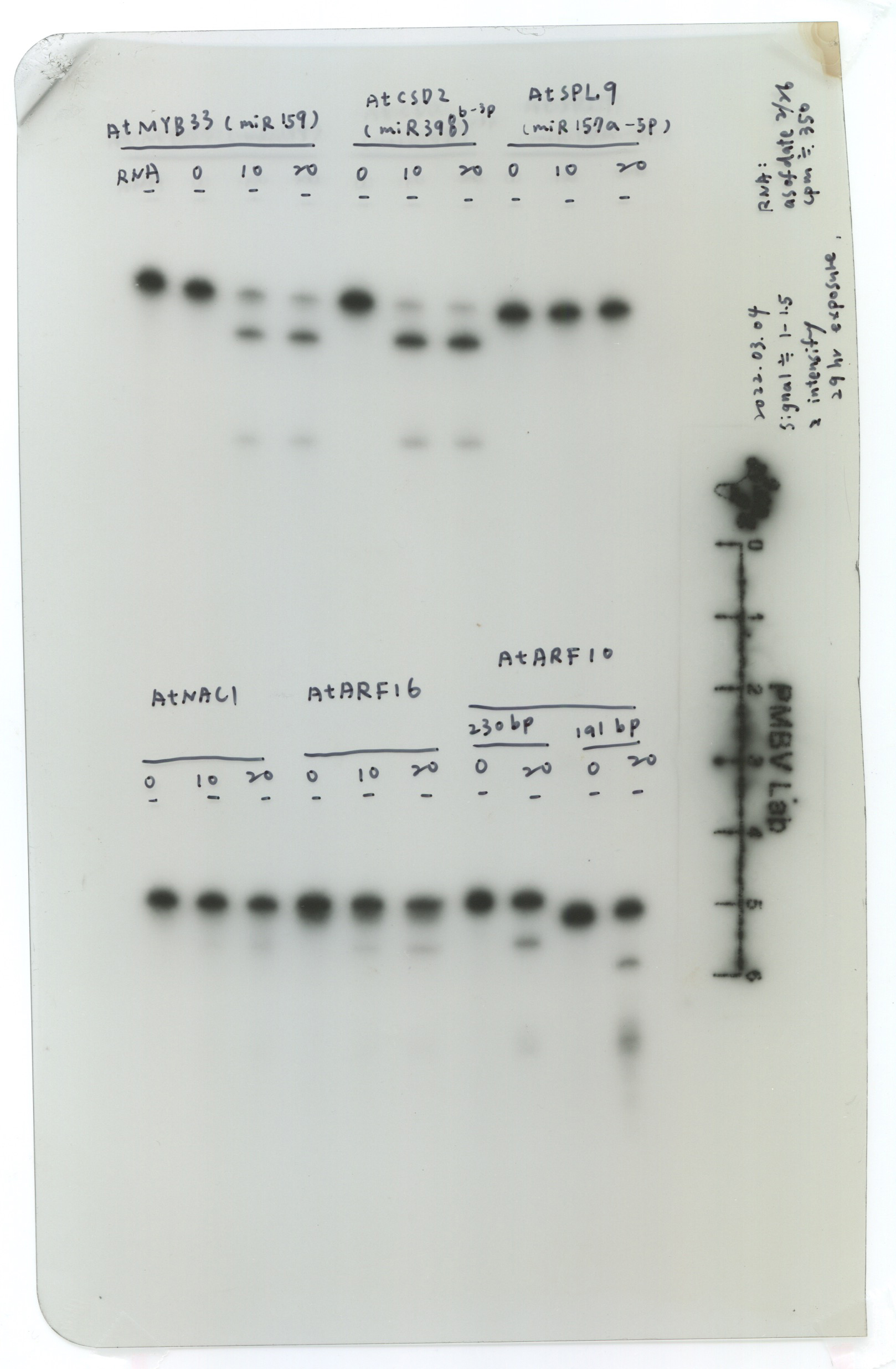
Next, we also examined different RNA substrates, including CSD2-230 (miR398 target), NAC1-230 (miR164 target), ARF16-230 (miR160 target), and ARF10-230 (miR160 targets) (Fig. 2g). All RNA substrates have different levels of cleavage efficiency, ex. MYB33-230 and CSD2-230 have higher cleavage efficiency than NAC1-230, ARF16-230, and ARF10-230 (Fig. 2h, panel i). We designed two different lengths of the 3'-end of ARF10 substrates, including ARF10-230 (230-nt) and ARF10-191 (191-nt) (Fig. 2g). The data indicated that ARF10-191 has better cleavage efficiency than ARF10-230 (Fig. 2h, panel ii). Thus, we assumed that the secondary structure of the RNA substrate and abundance of miRNA in AGO1 might affect the cleavage efficiency.
Evaluating endogenous RISC activity in P1/HC-Pro plants
Our previous study generated three viral species of P1/HC-Pro plants, including P1/HC-ProTu, P1/HC-ProZy, P1/HC-ProTe, and P1/HC-ProTu−K plants (Fig. 3a) (Kung et al., 2014; Hu et al., 2020). The P1/HC-ProTu, P1/HC-ProZy, and P1/HC-ProTe plants showed identical elliptical cotyledons (Fig. 3a, arrowheads). However, the cotyledons of P1/HC-ProTu−K plants were round in shape similar to those of Col-0 (Fig. 3a). We applied next-generation sequencing (NGS) to obtain the genomic sequences from these P1/HC-Pro plants. We also developed T-DNA finder software to identify the T-DNA insertion based on the genomic sequence profiles. Two T-DNAs were identified on chromosome 1, including P1/HC-ProTe (16,780,881 bp) and P1/HC-ProTu (28,814,559 bp) (Fig. 3b). In contrast, the P1/HC-ProTu−K is inserted on chromosome 3 (21,554,188 bp), whereas the P1/HC-ProZy is inserted on chromosome 4 (10,075,981 bp) (Fig. 3b). According to the TAIR database, the several critical RNA silencing component genes, e.g., AGO1, DCL1, and ATG8a, were also labeled on diagrammatic chromosomes with these T-DNA insertions, which is helpful for generating relative mutations in these P1/HC-Pro plants by crossing or gene editing (Fig. 3b).
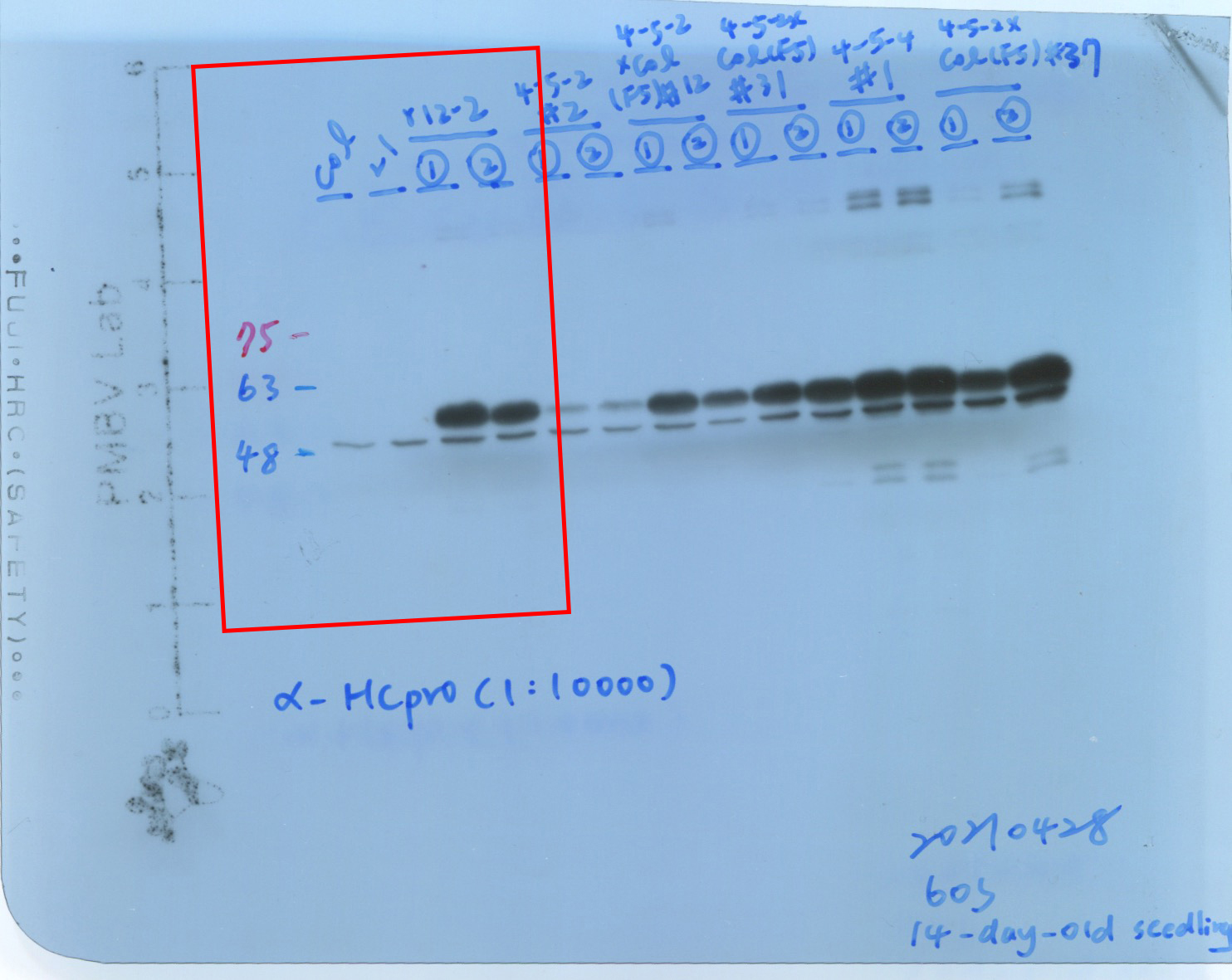
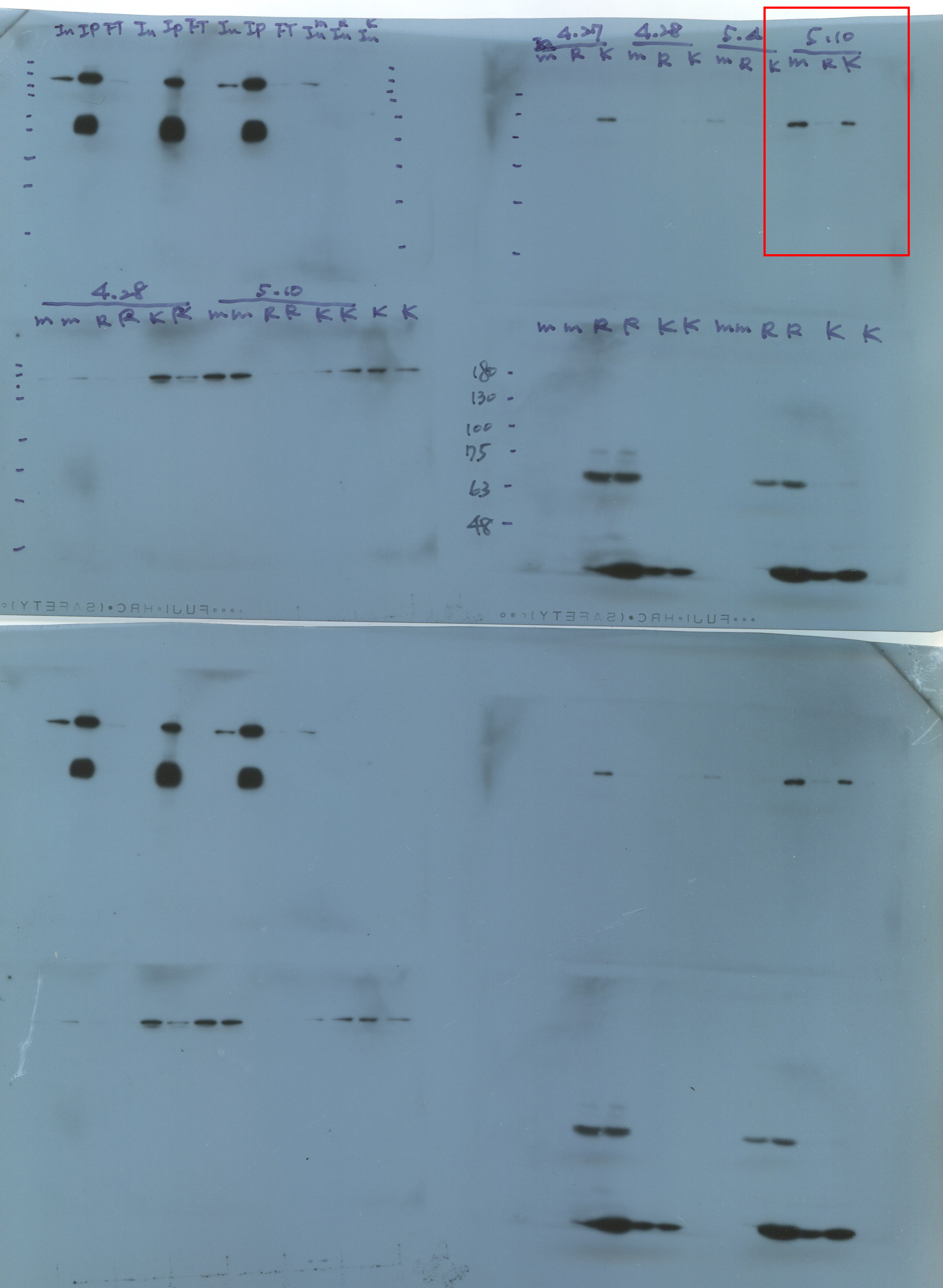
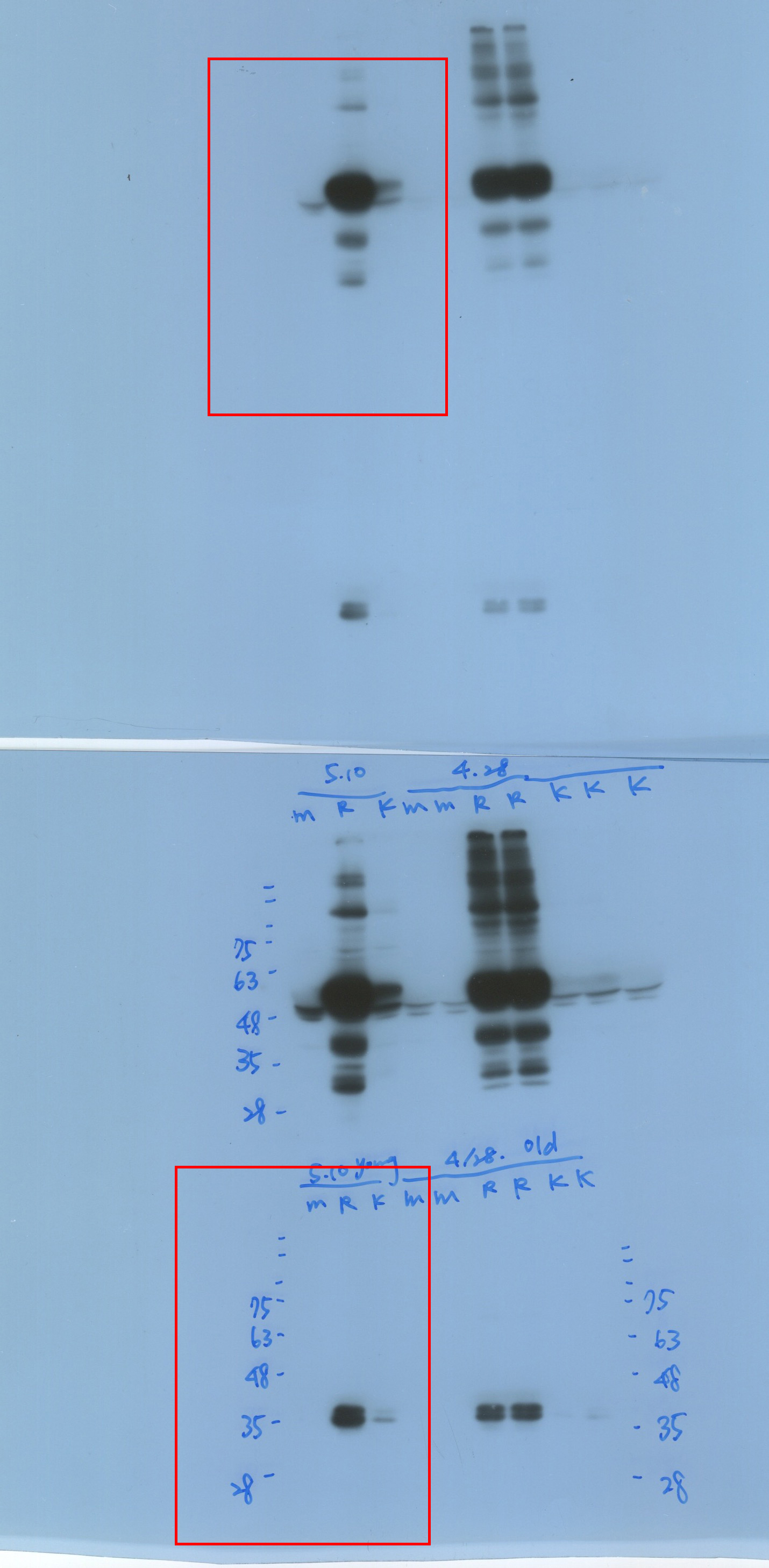
We performed western blotting to evaluate the endogenous AGO1 levels in various transgenic plants, including the L1 line and P1/HC-ProTu plants (Fig. 3c, panel i). Notably, the L1 line is a b-glucuronidase (GUS)-transgenic Arabidopsis, which underwent sense-posttranscriptional gene silencing and the T-DNA is inserted in chromosome 3 (2,350,388 bp) (Fig. 3b) (Elmayan et al., 1998), and the P1/HC-Pro plant was constructed on an L1 line background (Kung et al., 2014). The Col-0 and L1 lines showed identical endogenous AGO1 amounts, whereas P1/HC-ProTu plants showed lower levels of AGO1 while HC-Pro was presented (Fig. 3c, panel i).
Moreover, Hu et al. (2020) demonstrated that various P1/HC-Pro plants have different AGO1 levels in vivo. Indeed, P1/HC-ProTu plants had 0.5-fold higher AGO1 levels than Col-0 plants, whereas P1/HC-ProTu−K plants had 0.7-fold higher AGO1 levels, suggesting that HC-ProTu−K still has partial ability to trigger AGO1 degradation (Fig. 3c, panel ii). However, P1/HC-ProZy and P1/HC-ProTe plants have 0.7- to 0.9-fold AGO1 levels, suggesting that various species of HC-Pros have different AGO1 degradation efficiencies (Fig. 3c, panel ii). Therefore, we aimed to understand whether the endogenous AGO1 activity could be affected by VSR through in vitro RISC evaluation.
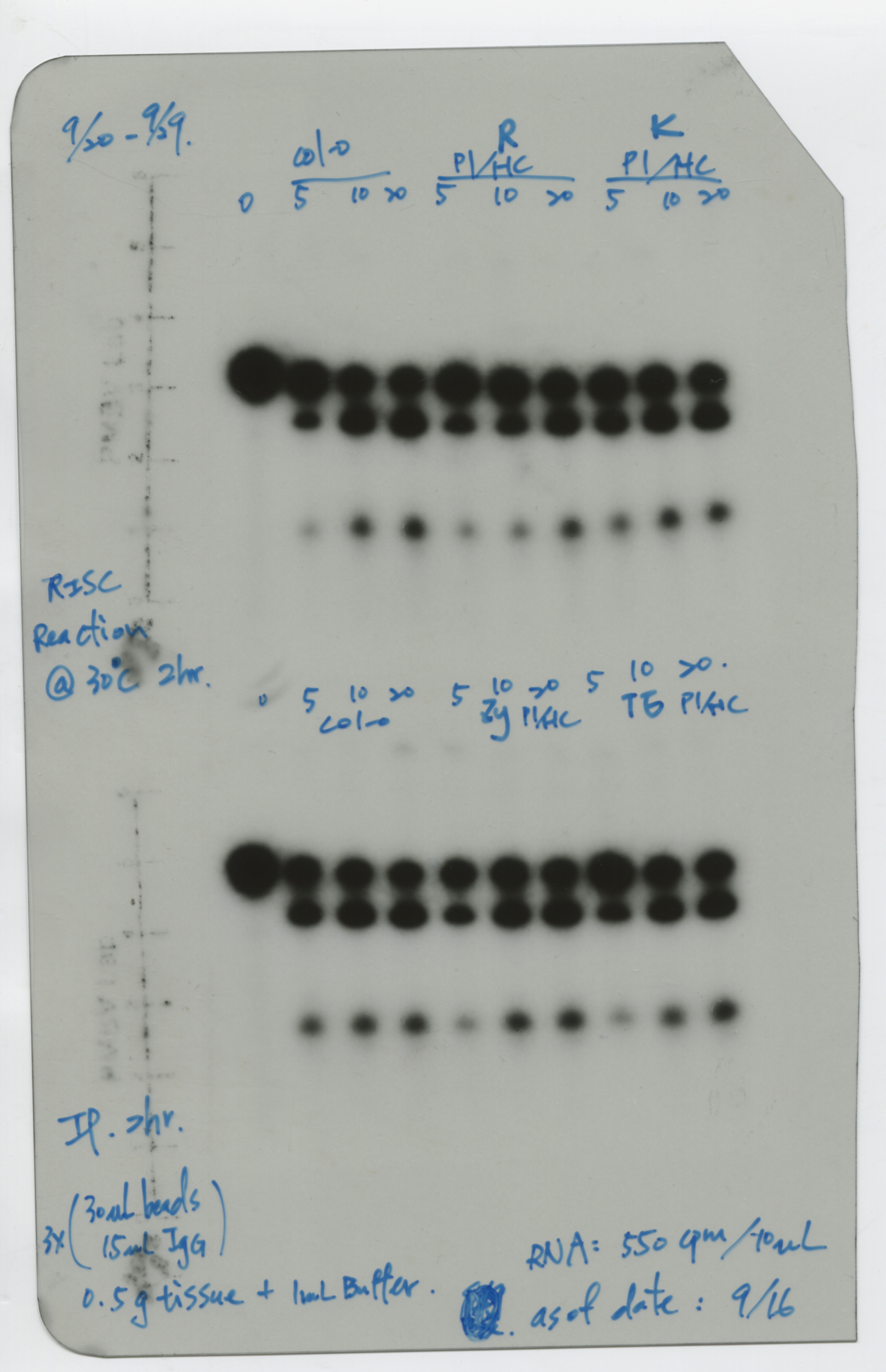
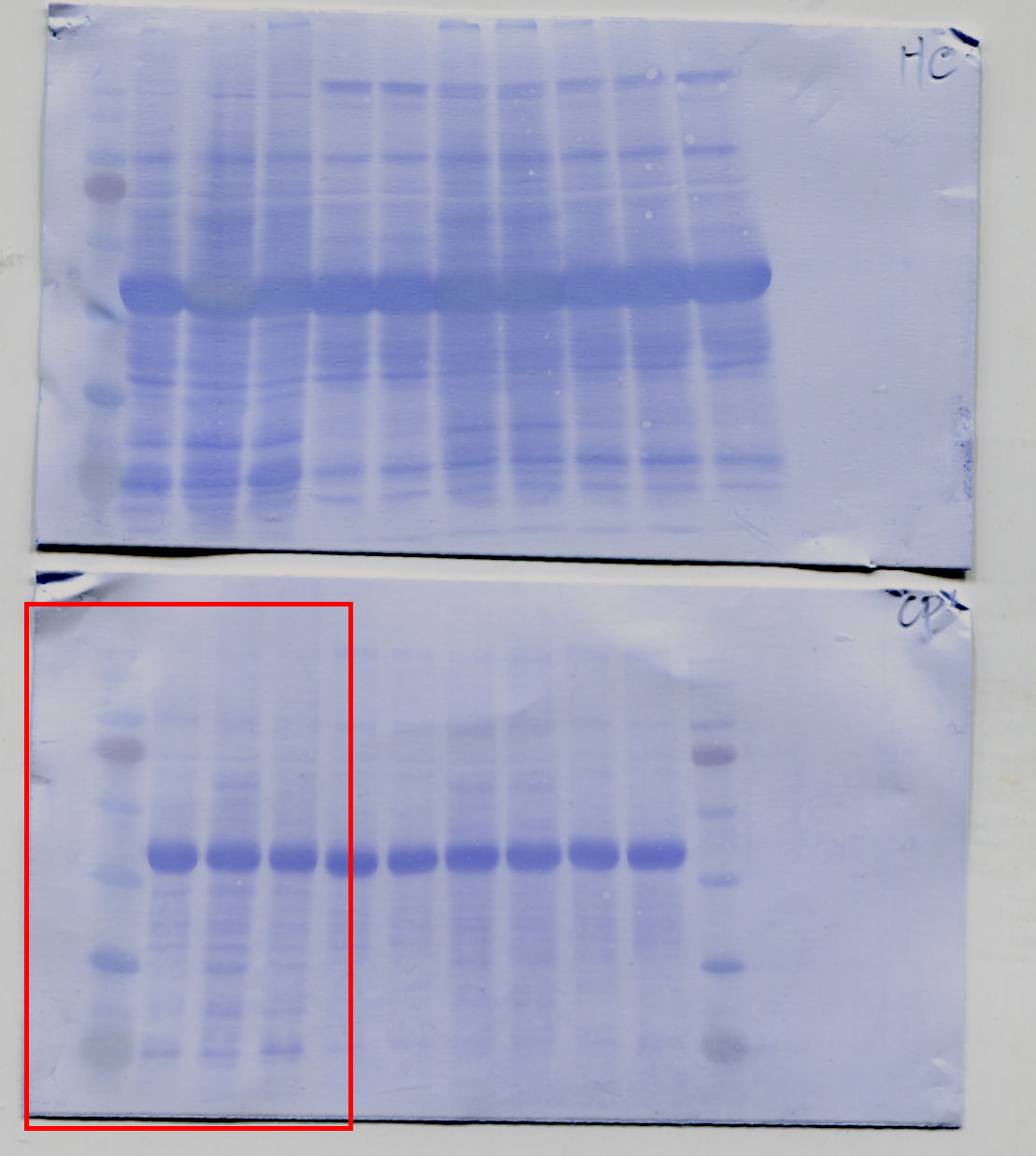
We purified comparable amounts of AGO1-IP from the various transgenic plants for in vitro RISC assays to evaluate the status of AGO1 activity with MYB33_230 RNA substrates. The AGO1-IP from Col-0 seedlings exhibited 20% and 29% normalized cleavage efficiency in 10 ng and 20 ng AGO1-IP, indicating that the cleavage is dose-dependent (Fig. 3d and e). In contrast, while the AGO1-IP from the P1/HC-ProTu plants still had RISC activity, the activity was reduced to 6% and 13% in 10 ng and 20 ng AGO1-IP, respectively, in comparable AGO1-IP amounts, suggesting interference with RISC regulation (Fig. 3d and e). Interestingly, we did not observe that AGO1-IP from P1/HC-ProZy and P1/HC-ProTe plants had different RISC activity compared with the AGO1-IP from Col-0 plants (Fig. 3e).
Furthermore, Wei et al. (2022) and Shang (2020) demonstrated that P1/HC-ProTu/atg8age plants, which have an ATG8a knockout mutant, have recovered AGO1 levels. Surprisingly, the normalized in vitro RISC activity returned to 20% and 39% efficiency at 10 ng and 20 ng, respectively, in P1/HC-ProTu/atg8age plants, suggesting restoration of RISC regulation (Fig. 3d and e). In summary, HC-ProTu triggers AGO1 degradation and inhibits RISC cleavage activity. However, HC-ProZy and HC-ProTe did not interfere with RISC activity.
TuMV-infected Arabidopsis triggers endogenous AGO1 degradation
Next, we performed endogenous AGO1 evaluation on TuMV-infected Col-0 plants (Fig. 4a). We evaluated the endogenous AGO1 levels in the TuGR- and TuGK-infected Col-0 plants. TuGR is the severe strain of TuMV that causes yellow mosaic symptoms on the leaves and up-curing leaves at the systemic leaves (Fig. 4a). Similar to P1/HC-ProTu plants, TuGR-infected Col-0 plants also showed lower amounts of endogenous AGO1 than mock or TuGK-infected Col-0 plants (Fig. 4b). A high abundance of HC-Pro and coat protein (CP) was detected in TuGR-infected plants, whereas lower amounts of HC-Pro and CP were detected in TuGK-infected Col-0 plants, which demonstrated uneven HC-ProTu and HC-ProTu−K amounts in infected tissues. Kung et al. (2014) and Hu et al. (2020) demonstrated that miRNA regulation was suppressed in P1/HC-ProTu plants. Similarly, miRNA regulation suppression was also observed in TuGR-infected Col-0 plants. More than 2-fold higher levels of AGO1 (miR168), ARF16 (miR160), and MYB33 (miR159) were also detected in TuGR-infected Col-0 plants (Fig. 4c). In contrast, TuGK infection caused ARF16 accumulation at approximately 1.5-fold, whereas RNA levels for AGO1 and MYB33 were similar to those of the mock treatment (Fig. 4c). These data indicated that miRNA regulation in TuGR-infected plants was also suppressed.
TuMV infection inhibits the RISC activity
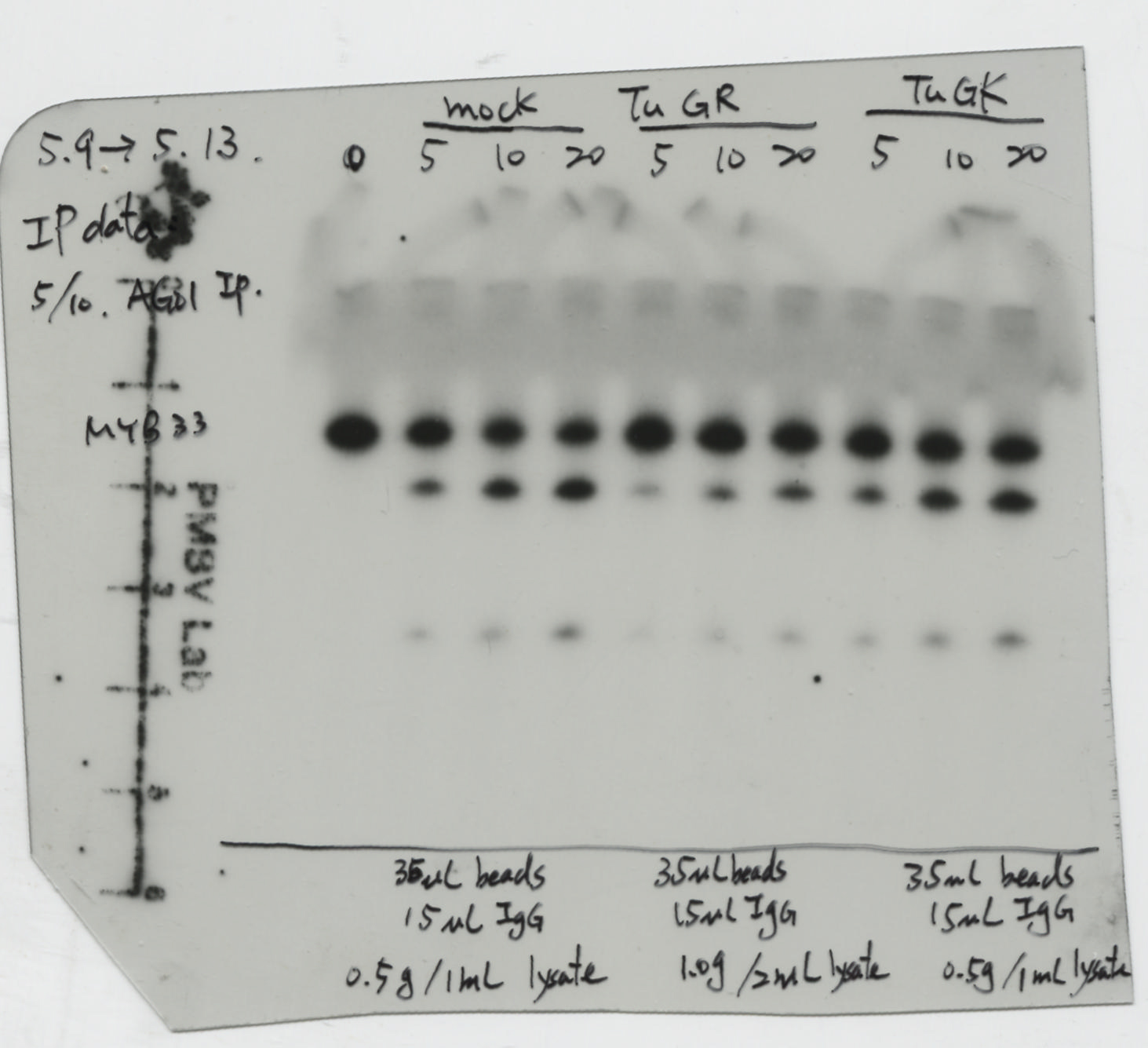
Next, we evaluated whether RISC activity might be affected during TuMV infection (Fig. 5a). We collected compatible amounts of AGO1-IP from mock, TuGR-infected, and TuGK-infected plants (Fig. 5b). The RISC assay demonstrated that the MYB33-230 substrate has low cleavage efficiency compared to mock or TuGK-infected plants (Fig. 5c). Figure 5c shows that 20 ng AGO1-IP from mock had 50.7% RISC activity, whereas 20 ng AGO1-IP from TuGR-infected and TuGK-infected plants had 38.5% and 43.9% RISC activity, respectively (Fig. 5a and c). These data demonstrated that severe TuMV infection inhibits RISC activity, which is consistent with the RISC results in P1/HC-ProTu plants.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}