Carbapenems and colistin are the last-resort antimicrobials against MDR organisms. The use of antibiotics is greatly challenged by NDM-mediated carbapenem resistance and MCR-mediated colistin resistance. According to V.M. de Carvalho Hessel Dias et al., the rate of carbapenem-resistant Acinetobacter baumannii (CRAB) per 1000 patient–days increased in 2020 and had a strong positive correlation with the incidence density of COVID-19, and CRKP displayed a gradual increase during the observation time [22]. Previous studies have shown that E. coli is the main reservoir for the NDM-5-IncX3 plasmid and mcr-1-IncHI2 plasmid. Herein we report coexistence of the NDM-5-IncX3 plasmid and mcr-1-IncHI2 plasmid in one strain of Klebsiella pneumoniae.
In our work, the blaNDM-5 genetic context is “tnpA-IS3000-IS30-IS5-blaNDM-5-bleMBL-trpF”, in line with the genetic context of plasmid pK516-NDM5 and plasmid pECNDM101. The good news is that IncX3 belongs to a narrow host plasmid [23]. Although it can carry the blaNDM-5 gene for conjugative transfer between different species, it has not been reported that it can cause outbreaks.
In addition, the existence of ISApl1, which is involved in the transposition of mcr-1, has been demonstrated. Our mcr-1 gene, located on plasmid pAN65-1, was linked to ISAPl1 and PAP2, corroborating previous studies [11, 24]. Intriguingly, the NFYY0065 chromosome also harbored the mgrB gene; mutation of the mgrB gene (usually point mutations caused by IS (insertion sequences)) will upregulate the expression of the PhoP/PhoQ system, resulting in colistin resistance [25–26]. Through our analysis, no IS were inserted in upstream or downstream of the mgrB gene, and the identity of the mgrB gene reached 100% with the genes in the NCBI database, which further confirms that the colistin resistance of the NFYY0065 strain is obtained from the product encoded by the mcr-1 gene on pAN65-1. In addition, pAN65-1 seems to comprise many other determinants, such as the IncHI2 plasmid [24]. IncX3 is a narrow-host-range plasmid; it plays an important role in the dissemination of blaNDM. Co-transfer of blaNDM-5 and mcr-1 by an IncX3–X4 hybrid plasmid in E. coli was reported by Sun Jian et al. [27]. In this study, IncHI2 and IncX3 type plasmids belonged to different incompatibility groups of plasmids; they can coexist stably in CRKP and have the ability of autonomous replication and horizontal transfer, and resistance genes on these two plasmids are transferred between different species of bacteria, thus forming a potential threat of outbreaks.
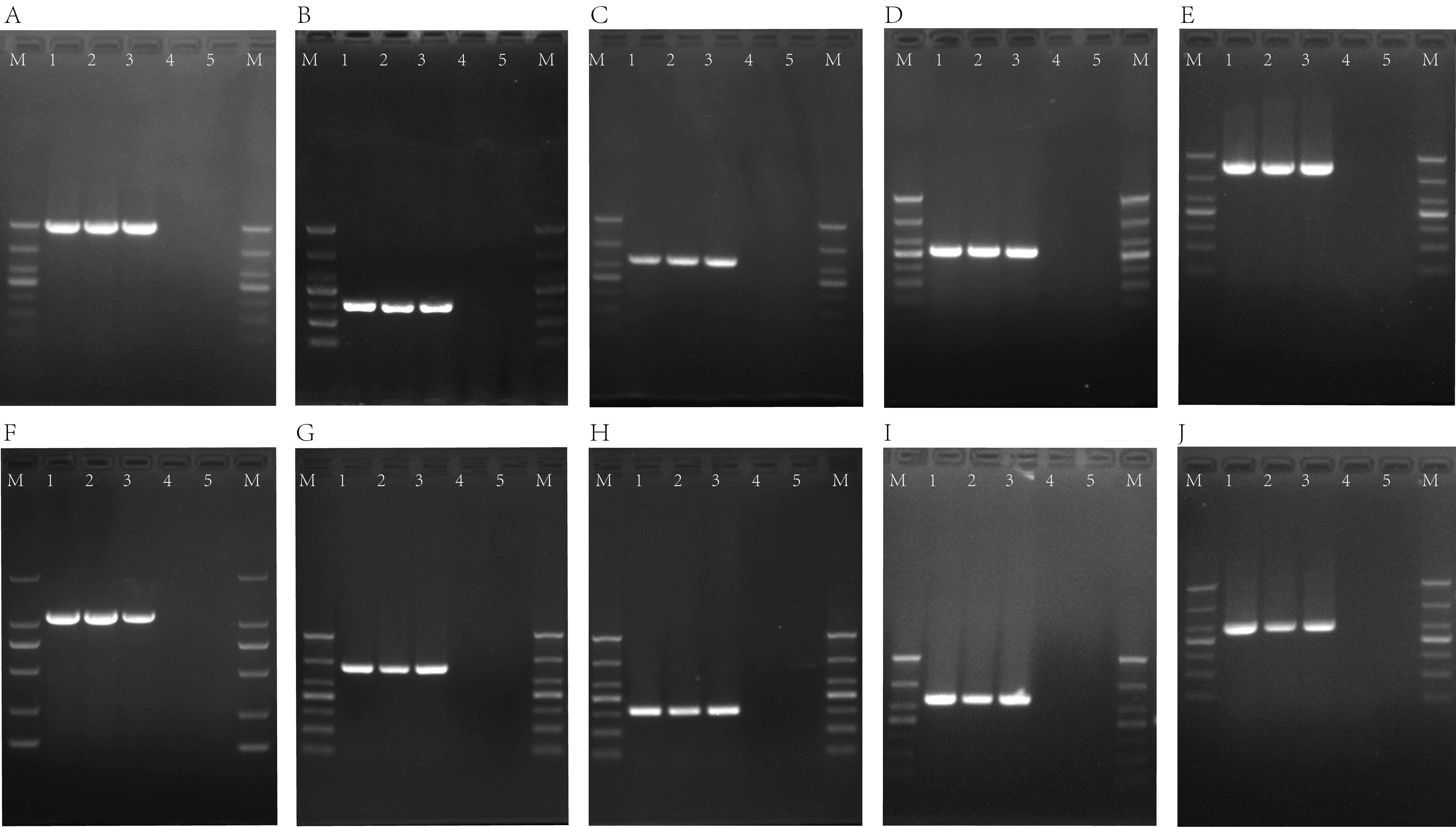
OMVs can be produced by growing cells of GNB. However, not all GNBs are able to transfer resistance determinants [28]. According to previous reports, OMV-mediated carbapenemase transfer occurs only with the following enzymes: NDM-1, OXA-24, and OXA-58 [15–16, 28].
This is the first report of NDM-5 carbapenemase transfer to susceptible bacteria mediated by OMVs released by CRKP. Unlike IMP, VIM, and SPM, which are periplasmic proteins, NDM is a lipidated protein that is anchored to the outer membrane of GNB, which facilitates the secretion of NDM into vesicles [29].
Previous studies have shown that additional OMVs may confer resistance to colistin in E. coli and P. syringae [17]. Li Xue et al. have suggested that mcr-producing OMVs reduce the protection in colistin-sensitive E. coli [30]; the mechanism of colistin resistance in Li’s research is due to the inability of colistin to bind intact outer membranes with lipid A-modification. We directly confirmed that OMVs are involved in the horizontal transfer of the mcr-1 gene located on plasmids, which means that OMVs could be a newly discovered HGT method to transfer mcr-1.
In hospital patients, the reservoir of KP is the gastrointestinal tract; in this study, NFYY0065 was isolated from a fecal sample, thus, suggesting the importance of stool management, given that feces is a highly important factor in the causation of nosocomial infections and that many classes of pathogens excreted in feces are able to initiate waterborne infections. Pathogenic bacteria “capture” resistance genes by conjugation transfer in the gut environment. Therefore, nosocomial infections caused by fecal contamination and water pollution should be strictly prevented.
In summary, this study provides the first report of a Klebsiella pneumoniae strain harboring both a blaNDM-5-IncX3 plasmid and a mcr-1-IncHI2 plasmid. For the first time we have demonstrated that OMVs can be used for the horizontal transfer of plasmids harboring blaNDM-5 and mcr-1 genes, which results in reduced susceptibility of the recipient strain to carbapenems and colistin. Although we found this phenomenon, the mechanism of vesicle-mediated horizontal transfer remains to be explored in more strains of bacteria.
{kind=link}