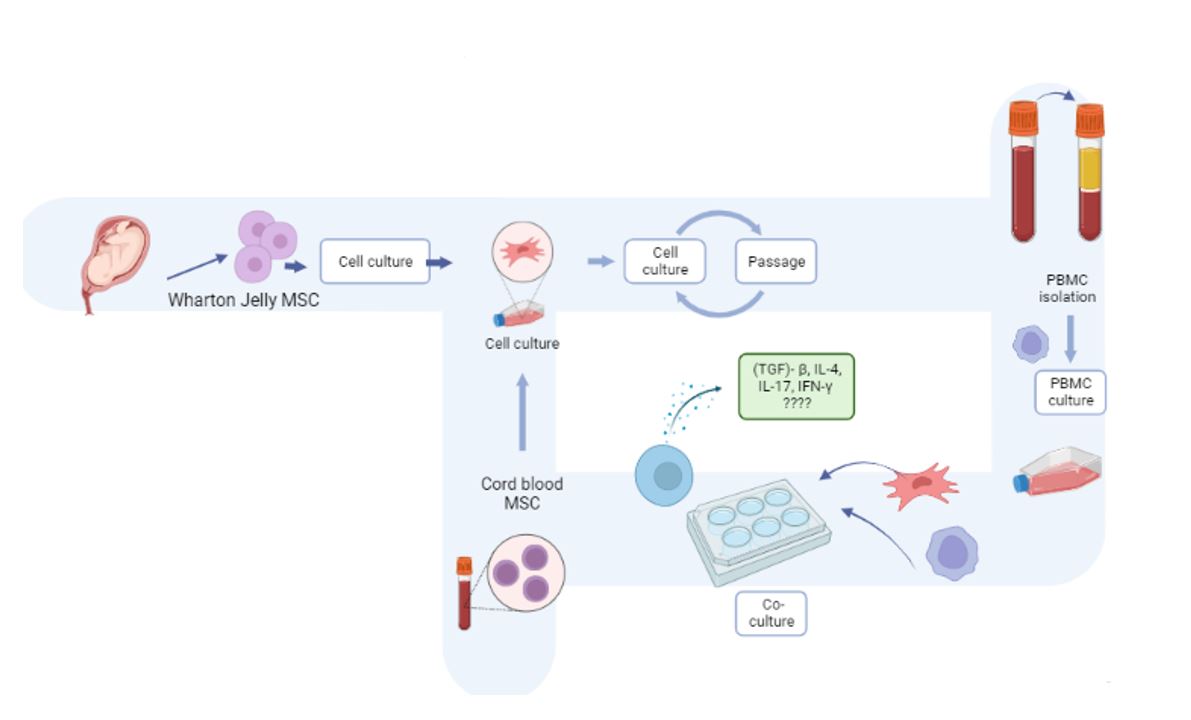
Mesenchymal stem cells are being studied as promising in the field of cellular therapy due to their self-renewal, immunomodulatory properties and multi-potential differentiation capacities. Although the in situ characteristics and biological functions of mesenchymal stem cells have not yet been elucidated, the in vitro obtained cells were shown to have the potential to differentiate into specific cell lineages. The studies have shown that MSCs have a different capacity to effect on immune system. They can modulate immune system interactions and suppress some inflammatory diseases. For some of autoimmune diseases, using MSCs as a cell based therapeutic are attracting attention. In many studies, the researchers have been focused on various mechanisms as secretion of cytokine and chemokine to detect anti-inflammatory and immunomodulatory effects of MSCs (29, 30).
The response of MSC to pro- and anti-inflammatory cytokines such as IFN-γ, IL-1, TNF, or IL-4 has been shown to be due to its mediators it secretes (TGF, IDO, PGE2) and with the conversion of CD4 + T helper cells into the Treg cell phenotype (31). T cells are key components, which are responsible for adaptive immune system and provides protection against malignancies, infections and autoimmune diseases. Several studies have been shown effect of MSCs on T cells through inhibition (32). Luz-Crawford et al. have reported that MSCs suppressed the secretion of IL-17, IFN-γ however induced the IL-10 production of the T cells by antagonising the differentiation of Th17, and Th1 cells and thus induced the Treg formation (33). Yang et al. have demonstrated that IFN-γ was significantly suppressed in the comparison of WJ-MSCs administered PBMCs and PBMCs stimulated with PHA without WJ-MSC in the examination of immune system stimulants detected in cells and supernatants, which were collected after co-culture of PBMCs with WJ-MSC (26). In our study, the IFN-γ level in the supernatant after co-culture was found to have decreased significantly compared with the STI-PBMCs that were not performed WJ-MSC at the end of the hour 24, and hour 72. For the determination of the intracellular cytokine, PBMCs were obtained from culture plates and studied, and for the supernatant, the culture fluid containing MSC and PBMC after co-culture was collected and studied. We suggest that MSCs inhibited the cytokine release out of the cell from PBMCs as the reason for the difference in the amount of intracellular cytokine and cytokine amount released out of the cell. The MSCs isolated from Wharton’s Jelly inhibited the intracellular release of the pro-inflammatory cytokine IFN-γ at the end of hour 72 in the 1/5 co-culture of WJ-MSC/PBMC. The separate investigation of the IFN-γ intracellular and supernatant values of MSCs isolated from the cord blood showed that the intracellular cytokine level significantly increased, and the cytokine level in supernatant significantly decreased for each group at hour 24 and hour 72 compared with the group that was not performed CB-MSC. The addition of CB-MSC on PBMC inhibited the IFN-γ cytokine values only in the supernatant.
The T cell subsets of Th17 cells in the immune response, express the chemokine receptor CCR6 and a transcription factor of RAR associated receptor T (RORyT) as a distinguishing factor of Th17 specific T cell in addition to the expression of IL-17A, IL-17F and IL-22 cytokines. Th17 cells, which are involved in the pathogenesis of various inflammatory and autoimmune diseases such as multiple sclerosis, systemic lupus erythromatosus, Type 1 diabetes, and rheumatoid arthritis, play role in insufficient or deficient T cell immune regulation. Although the mechanisms by which MSCs modulate the response of the Th17 cells subgroups, some studies have been reported to date but that is not enough to explain all mechanisms (34, 35, 36, 37). Mesenchymal stem cells can directly or indirectly inhibit Th17 differentiation and functions (38, 39). MSCs can inhibit Th17 differentiation by increasing the expression of CCL2, PD-1, IL-10, or SOCS3. Some studies show that MSCs may also promote Th17 proliferation (40, 41, 42). Guo et al. have shown that MSCs isolated from bone marrow promote IL-17 expression and Th17 cell differentiation (43). In our study, we also observed that WJ-MSC added at different rates on stimulated PBMC significantly increased the intracellular expression of IL-17 after co-culture at different times. As the co-existence times of PBMC and MSC increased, the intracellular cytokine expression decreased. However, the cord-derived MSC showed the opposite effect and the intracellular level of IL-17 cytokines increased higher by 1/5 ratio of the cell. The examination of the IL-17 cytokine levels in the supernatant at the end of MSC and PBMC co-culture showed significant inhibition in both WJ, and CB. Compared with the conducted studies, the level of IL-17 supernatant cytokines may be inhibited by MSC. On the other hand, further studies are needed for the intracellular cytokine level. It is suggested that the quality of MSCs, where they were isolated from, and the difference in their ratio to T cells may change their effect on cytokine levels.
Mesenchymal stem cells support the maturation, differentiation of Th2 cells and secretion of IL-4 from Th2 cells through the expression of IDO. Gieseke et al. have indicated that MSCs might directly suppressed the T cell proliferation (44, 45). MSCs have a pro-inflammatory enhancing effect and an anti-inflammatory suppressive effect on the immune system. Aggarwal et al. have found in their study evaluating the supernatant values after co-culture of the MSCs isolated from the bone marrow with the T cells isolated from the BMC by 1/10, that the IL-4 level significantly increased (46). In our study, the intracellular IL-4 ratio significantly increased in each group in the groups where CB-MSC was used. The level of IL-4 in the supernatant was significantly suppressed in the groups where both sources of MSC cells were used.
The main immunomodulatory cytokines of IL-10 and TGF-β are the cytokines produced by the MSCs. TGF-β is structurally secreted by MSCs, and its level can also be increased by inflammatory factors such as TNF-α and IFN-γ. TGF-β inhibits the expression of IL-2, MHC-II (major histocompatibility complex II) and T cells. Both Th1 differentiation and Th2 differentiation can be suppressed by TGF-β. In addition, the immunosuppressive effects of bone marrow-derived MSCs stimulated by TGF-β, IFN-γ and TNF-α were shown to have been eliminated by inhibiting the expression of iNOS and IDO (47, 48). Yang et al. have shown that WJ-MSCs and PHA-PBMC stimulated by co-culture supernatant detected in collected PGE2, TGF-ß1 and IL-10 Treg immune system on examining stimuli, WJ-MSCs applied PBMC with WJ-MSCs without PBMC stimulated with PHA when compared to PGE2, TGF-ß1 and IL-10 have shown that greatly increases the number of Treg cells. Along with this finding, they emphasized that WJ-MSCs have an immunosuppressive effect by increasing the production of suppressive cytokines (26). Wang et al. have found that the Th1 and Th17 cells were suppressed, and the count of the Th2 and Treg cells increased after 5-day co-culture of 1/5 ratio of stimulated WJ-MSC/stimulated PBMC (49). In our study, the TGF-β cytokine level was determined by ELISA from the supernatant after co-culture in all experimental groups. The level of TGF-β1 cytokines in the supernatant showed an important and significant increase after 24-hour co-culture of WJ-MSC/PBMC by 1/5 ratio. The study of Wang et al. supports our study and the level of TGF-β cytokines secreted from Treg cells increased significantly by 1/5 cell ratio (49). We suggest that the reason for obtaining different results than other studies could be due to the use of stimulated MSCs, and co-cultures times were longer, was suppressed in our other groups.
Deuse et al. have emphasized that the use of higher ratio of MSC inhibits the T cell proliferation, and the use of MSC in smaller ratio contributes positively to T cell proliferation. They suggested that the ratios of PBMC and MSC in co-culture may have different immunomodulation effects by affecting T cell proliferation, and that cell-cell interaction is not essential in their study (35). In our study, we used 2 different ratios as 1/5 and 1/10 in direct MSC/PBMC co-culture, and different results were obtained at different ratios. The study of Krampera et al. also supports our study and they emphasized that a higher amount of MSC has a greater ability to suppress on T cell (50). The comparison of these two different ratios showed that the level of IL-17 cytokine was significantly increased both in intracellular cytokine and supernatant cytokine at the end of 24 hours and 72 hours at 1/10 cell ratio compared to the levels at 1/5 cell ratio. The levels of IL-4 intracellular cytokine and TGF-β cytokine in the supernatant significantly increased at the end of 72 hours at 1/10 cell ratio compared with the levels at 1/5 cell ratio. We suggest that administration of higher amount of MSC on PBMC increases cytokine production and secretion by increasing T cell proliferation. In our study, while the count of MSC remained constant, the PBMC count was proportionally increased. In co-cultures where MSC was relatively higher, it stimulated T cell proliferation, which led to higher cytokine expression and secretion.
The MSCs derived from adipose tissue and bone marrow are generally used in the studies of mesenchymal stem cells on the immune system. In this study, the use of Wharton's Jelly and cord blood as a source of MSC and the investigation of its effect on the immune system provided a different perspective to the literature. The effect of different sources of MSC on the immune system in the study was evaluated in different combinations in the form of using 2 different cell ratios of 1/5 and 1/10, and two different incubation periods of 24 hours and 72 hours. In terms of the separate evaluation, analysis, and comparison of these parameters, this study has an original value.
Despite all this promising research, the presence of ambiguous results in clinical studies with MSC, caused by the variability of MSC populations on the immune system hinders the progress of the studies. Therefore, the investigation of both intracellular and supernatant levels of pro-inflammatory and anti-inflammatory cytokines in our study has given a new perspective to this issue. There is no available previous study on how MSC affects the level of intracellular cytokines in the cells of the immune system; the study is original in this respect.
In the study, obtaining different supernatant and intracellular cytokine values suggests that MSCs may be effective in preventing the transport of cytokines expressed inside the cell to the outside of the cell. Detection of the mechanisms of the extracellular transport pathway of cytokines expressed in the cell with further studies will contribute to understanding the cause of the intracellular cytokine level difference with the supernatant.
The interaction of MSC and immune system cells initiates a complex process. In order to elucidate this complex process, the correct and appropriate selection of the source of MSC can be possible by specifically identifying the subgroups of cells belonging to the immune system, as well as in in vitro and in vivo studies.
{kind=link}