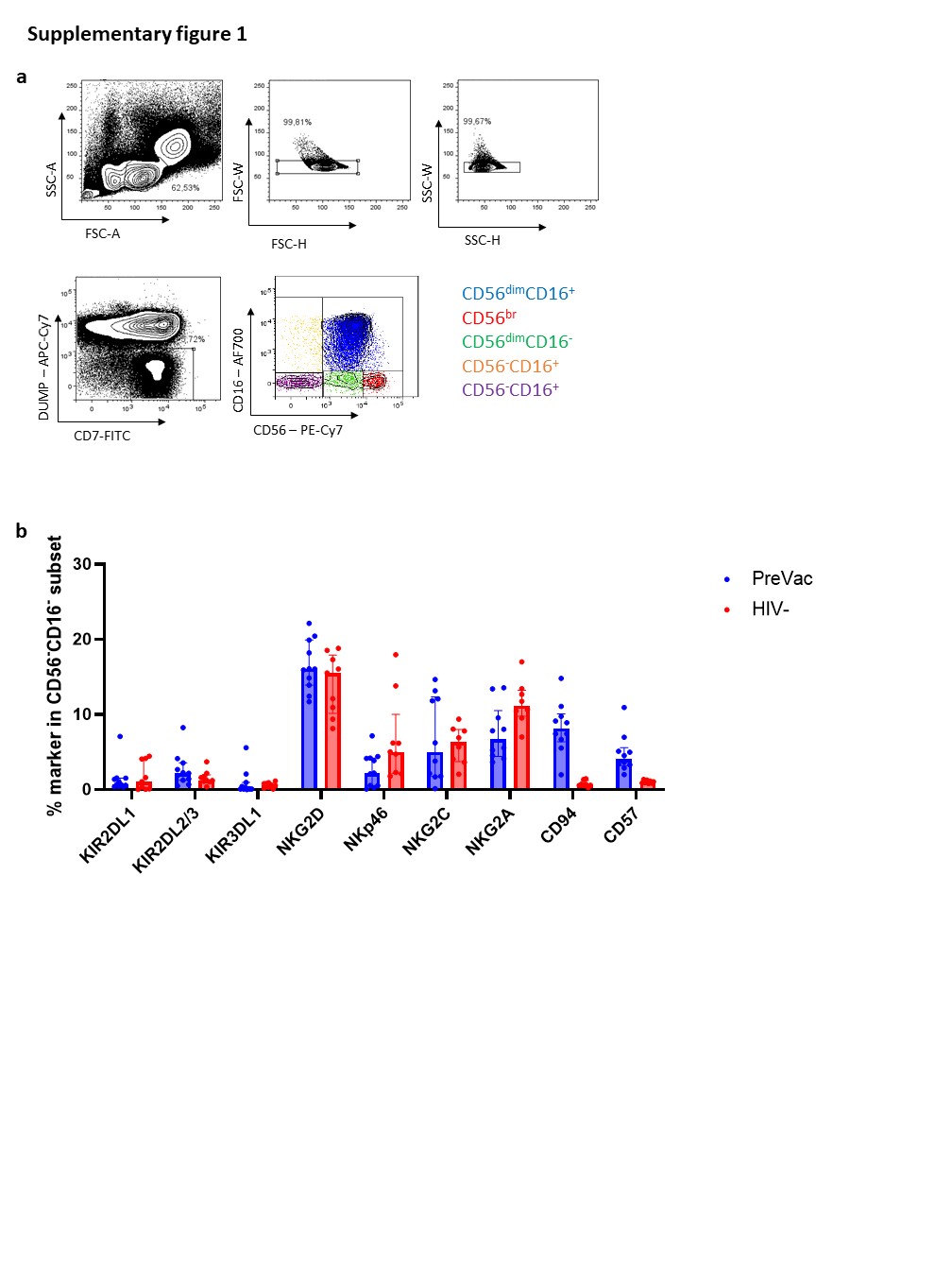
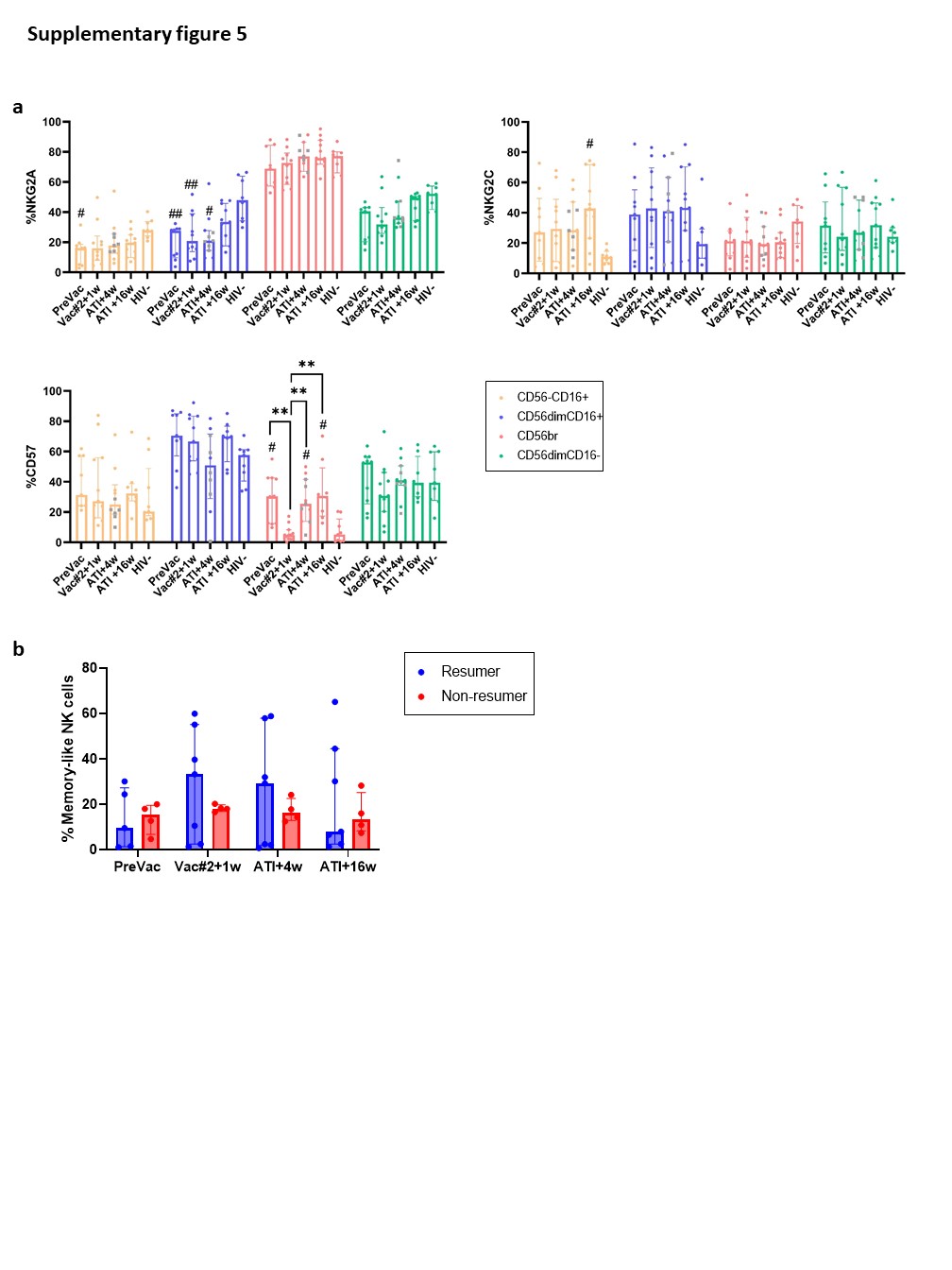
In this study, we investigated whether autologous DC-based immunotherapy affects NK-cell frequency, phenotype and functionality. Although we did not observe a difference in the frequency of total NK cells, we observed significant redistributions of NK-cell subsets upon DC-based vaccination and subsequent ATI. At Vac#2 + 1w, we identified an increase in the cytotoxic NK-cell subset concomitant with a decrease in the CD56dimCD16− subset. The decrease in CD56dimCD16− NK cells in peripheral blood might be explained by the migration of NK cells to the lymph nodes where interaction with DCs is likely taking place. Another possible explanation would be the maturation of CD56dimCD16− NK cells into CD56dimCD16+ NK cells33,34. The latter possibility is less likely since a trend towards reduced CD57 expression, which is an important maturation marker, was observed for all NK-cell subsets at Vac#2 + 1w compared to baseline. This is in line with data published by Di Nicola et al. who also reported a significant increase in cytotoxic NK cells upon autologous tumour-loaded DC vaccination against B-cell lymphoma35. Pena et al. performed a trial in which PLWH received three immunizations two weeks apart with monocyte-derived DCs pulsed with heat-inactivated autologous virus. NK cell frequency was studied at 1, 3, 16, 24 and 48 weeks after the first vaccination36. Results showed that the frequency of CD56dim NK cells was significantly decreased at week 24 but not at the other time points, whereas we found a significant early decrease of the CD56dimCD16− subset at Vac2 + 1w. Several studies reported a loss of CD56br NK cells and an increase in the dysfunctional CD56− population when PLWH are experiencing high viral loads, which is in line with our results24,37,38.
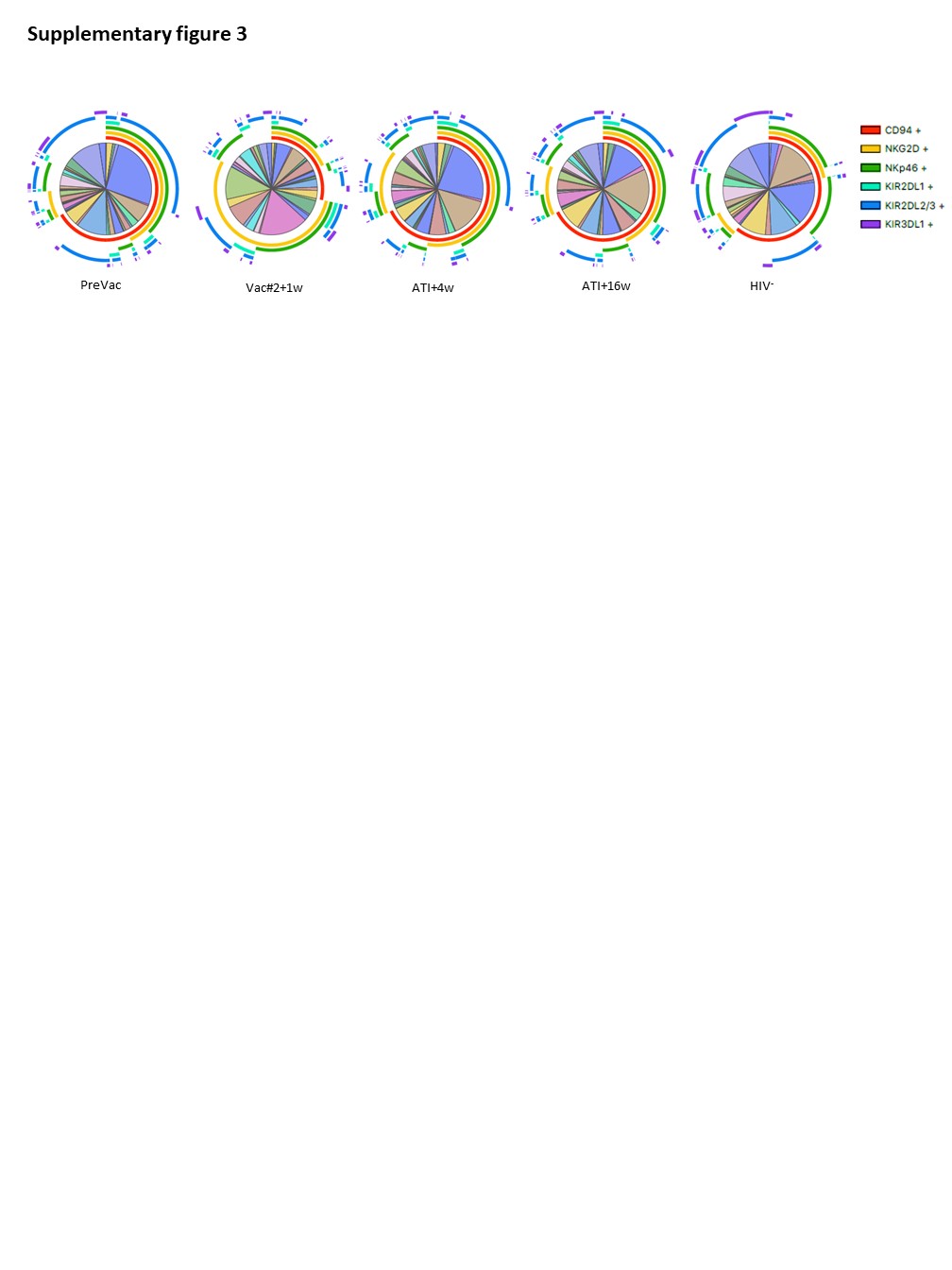
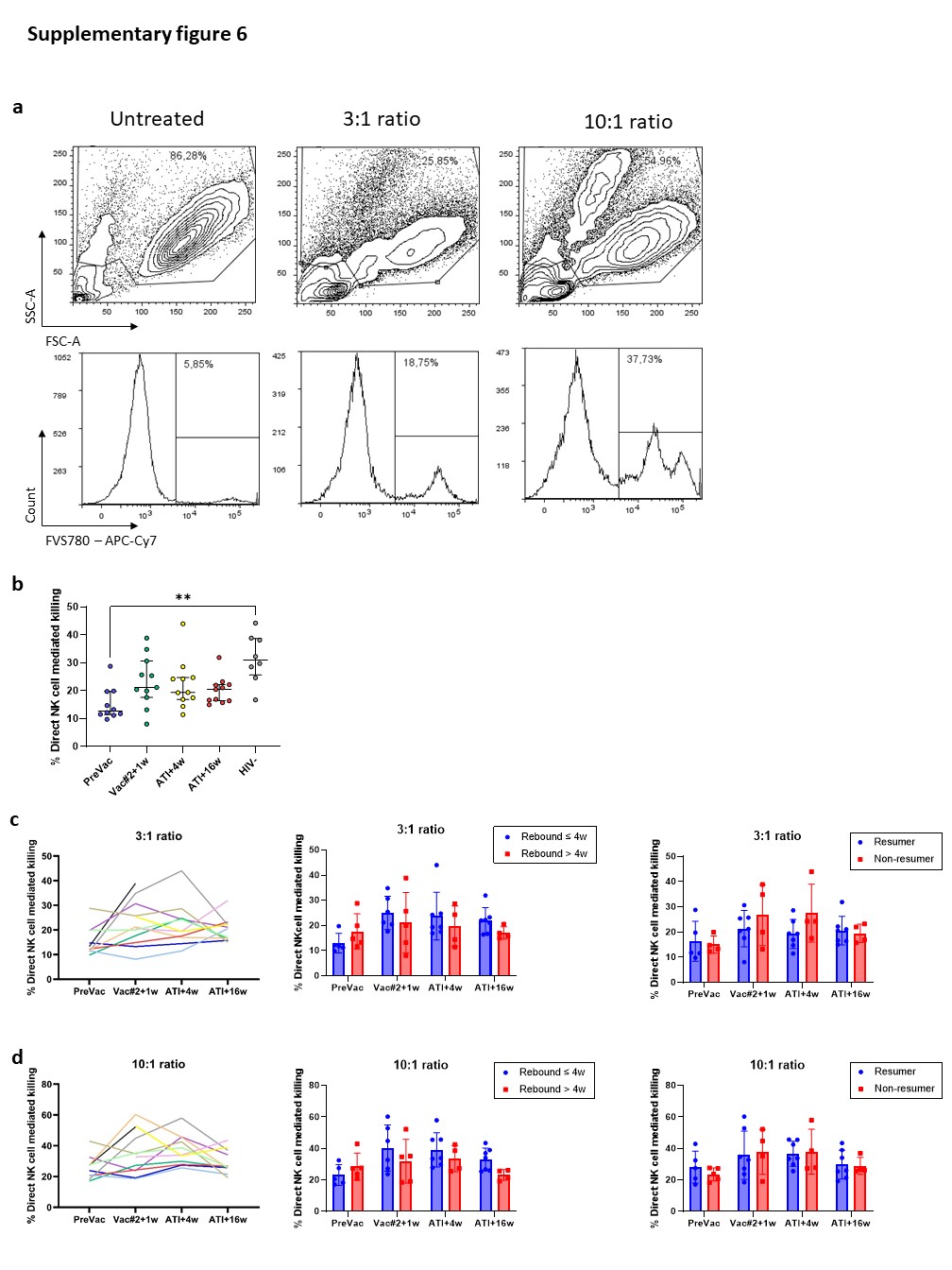
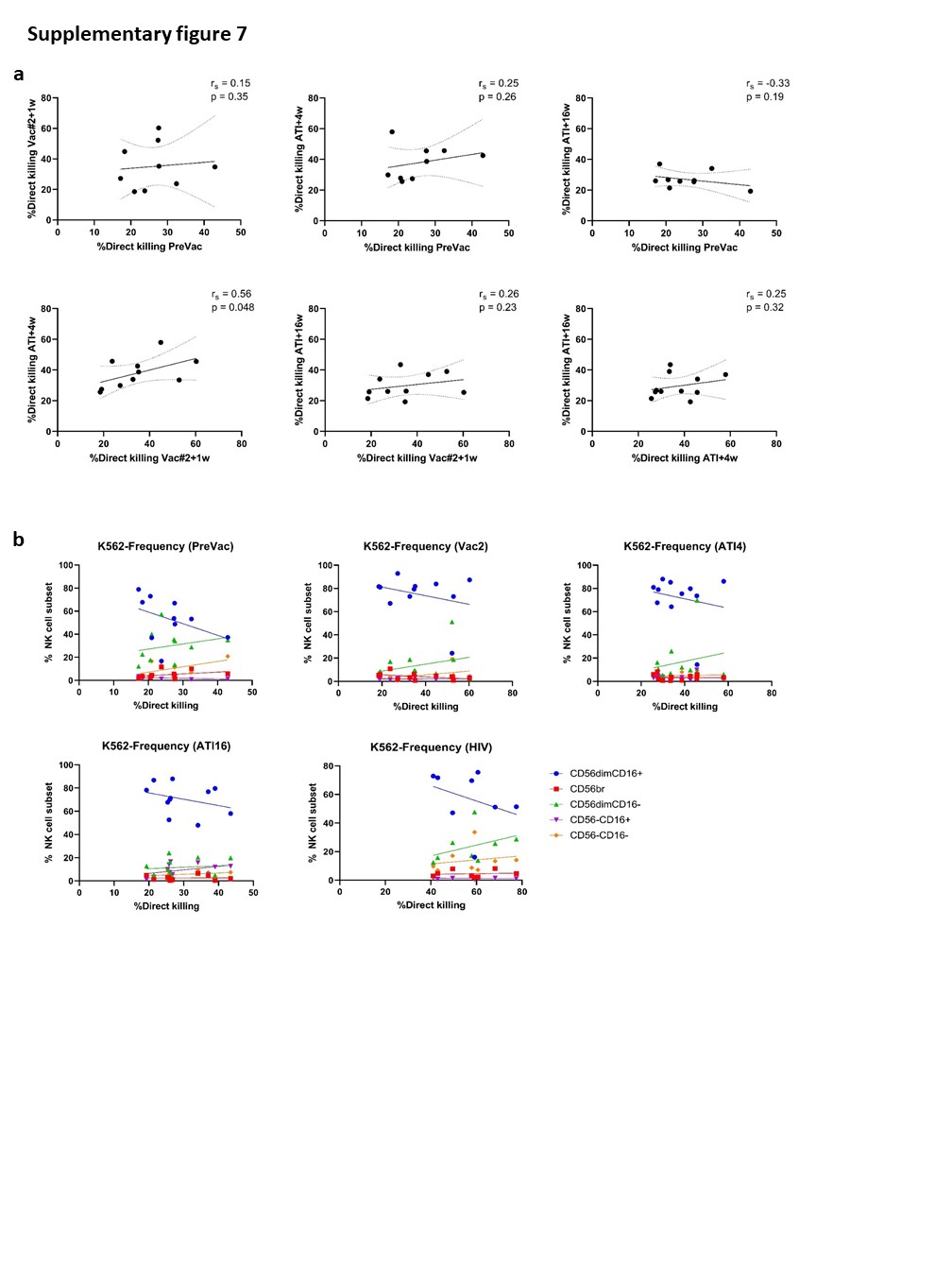
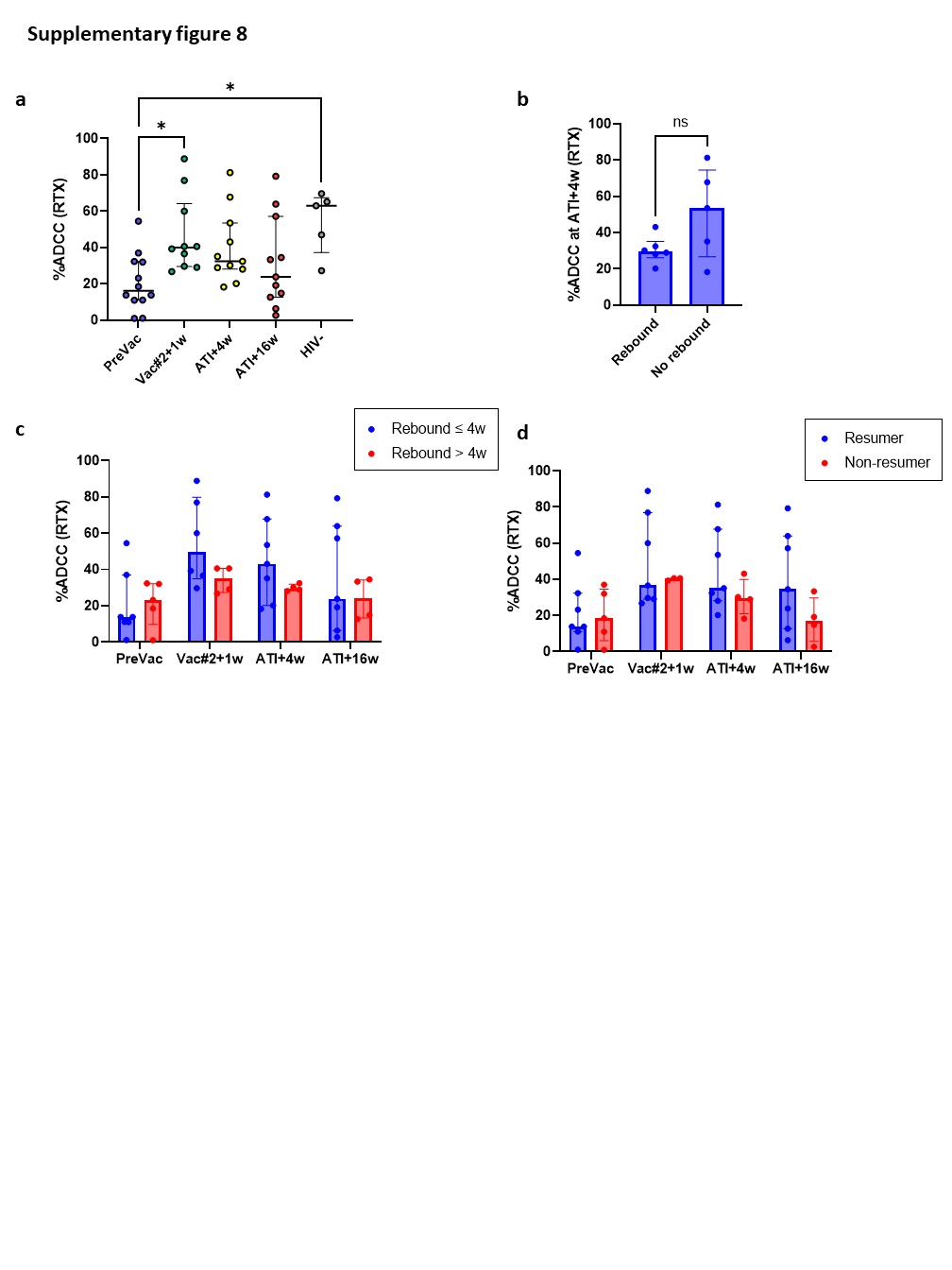
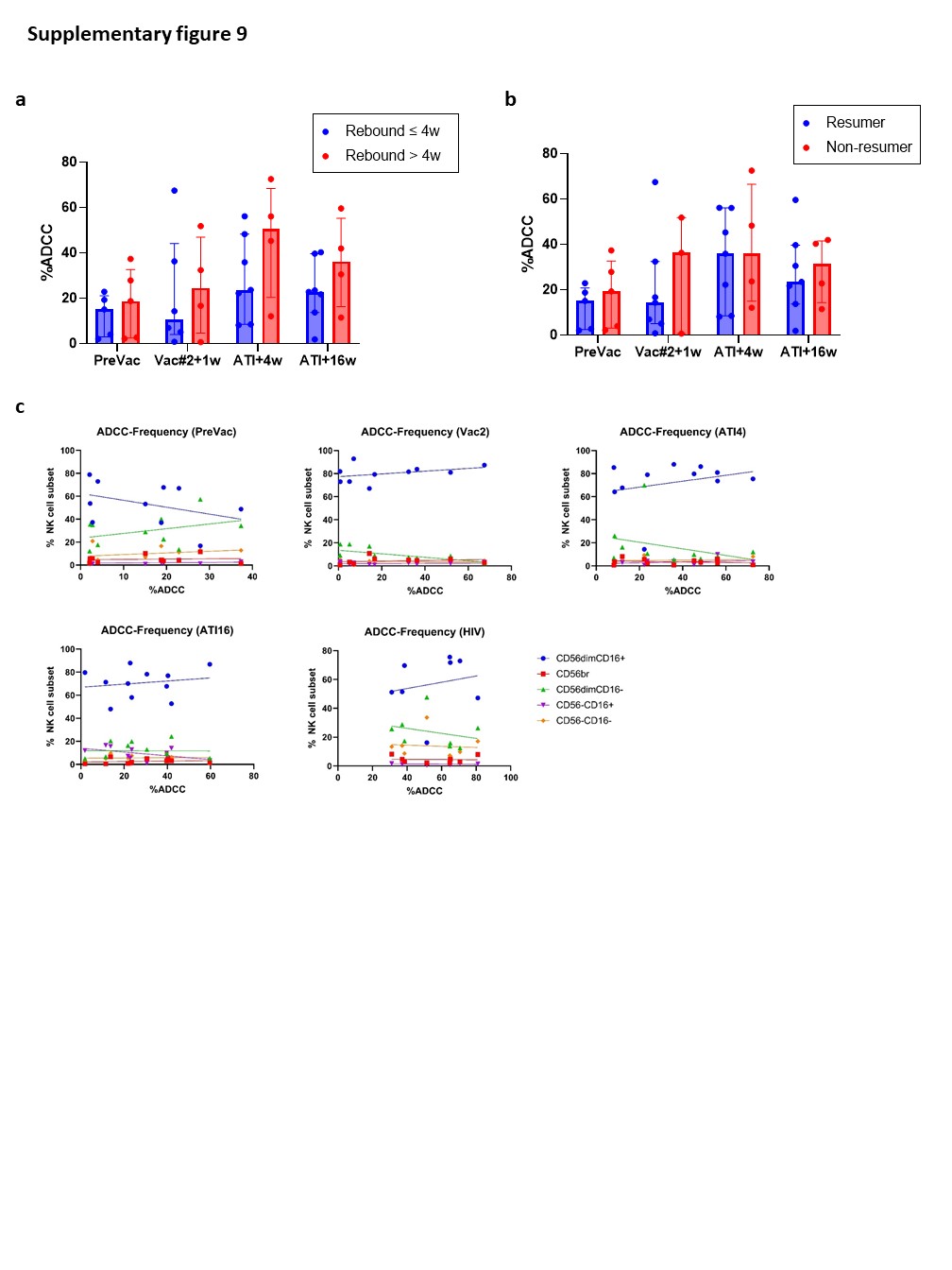
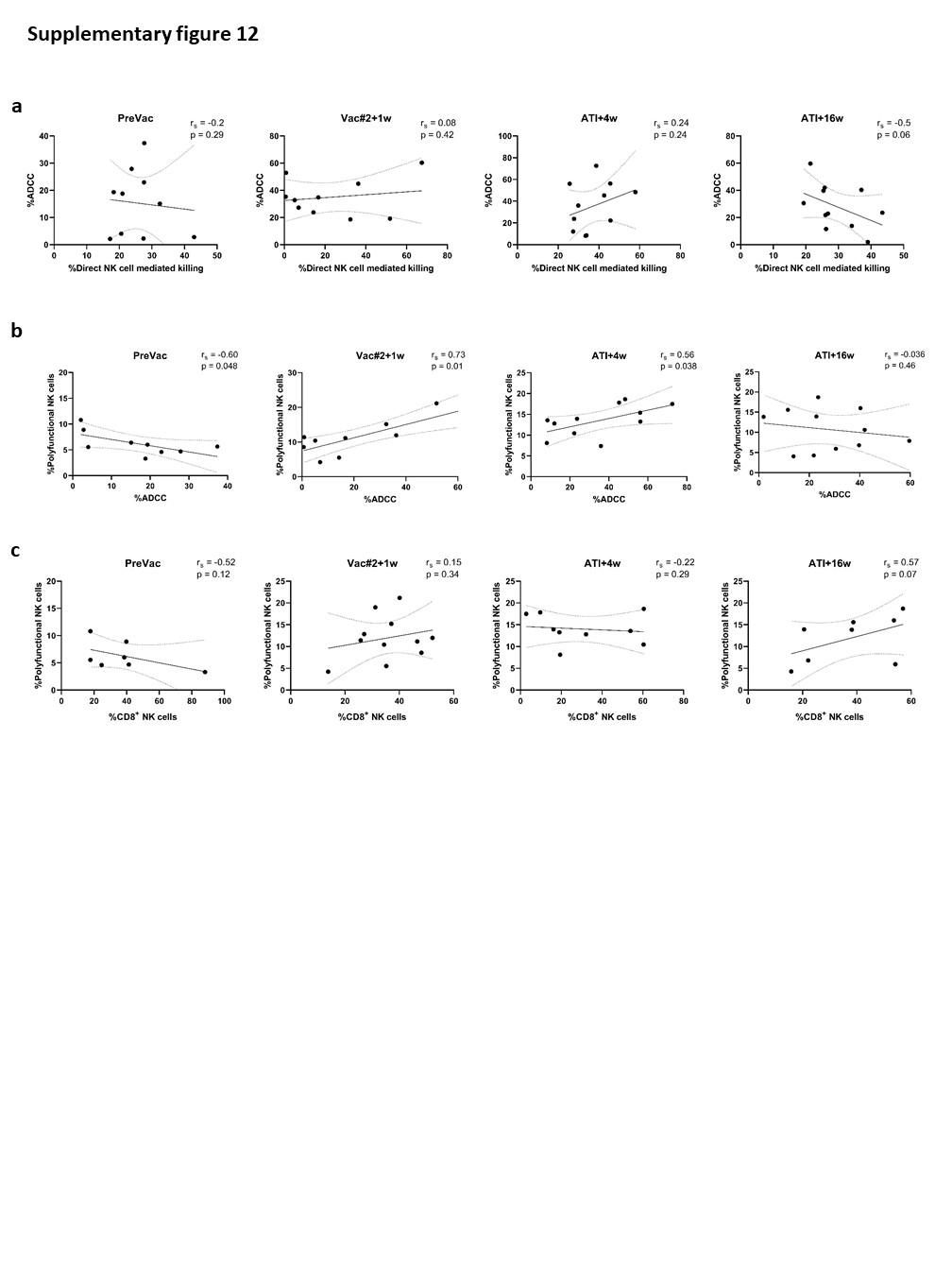
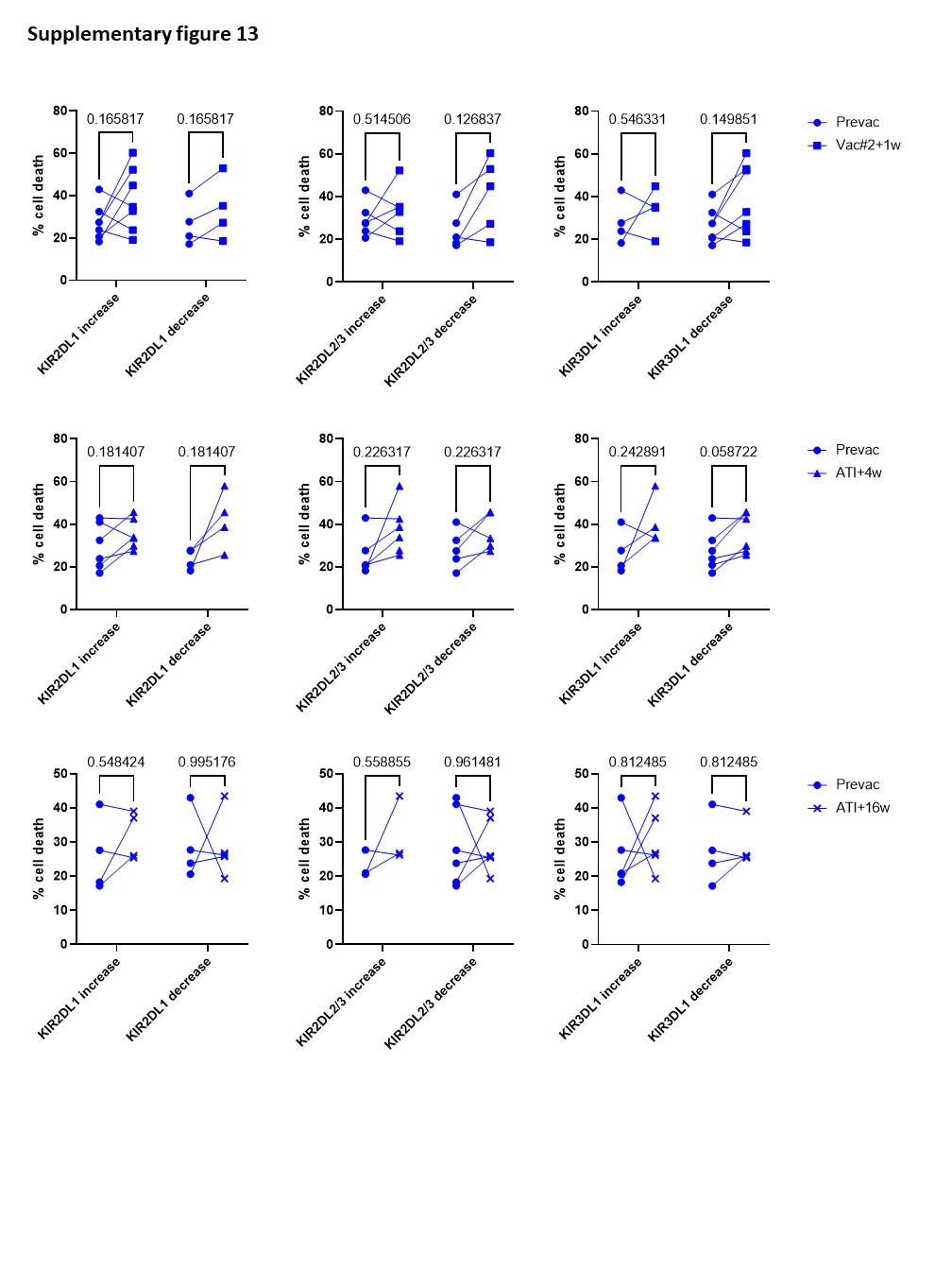
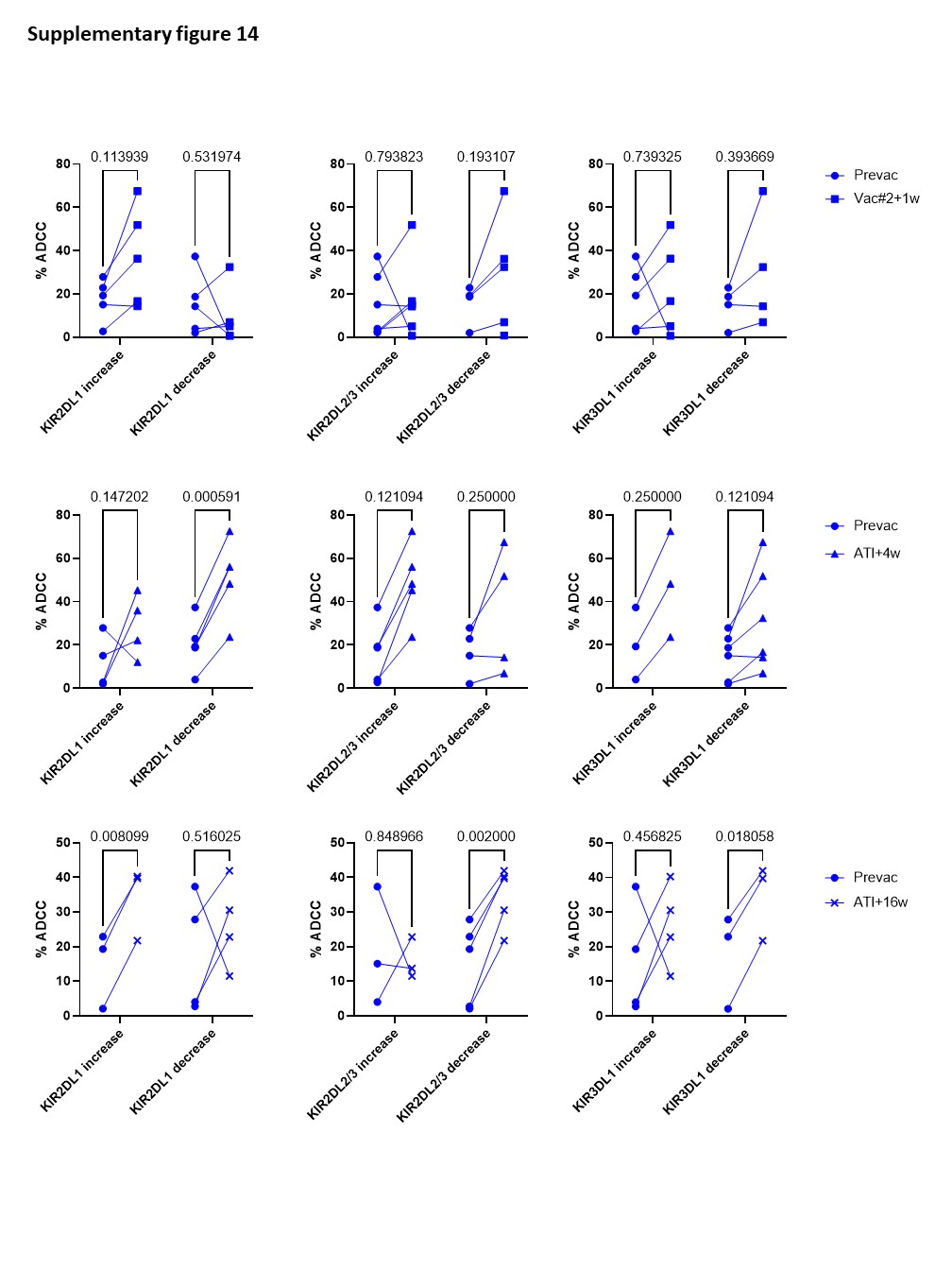
Although the frequency of cytotoxic NK cells significantly increased at Vac#2 + 1w, their expression of both activating and inhibitory receptors did not change. In contrast, based on KIR expression, the CD56dimCD16− subset showed a more educated phenotype. Kristensen et al. showed that educated NK cells are superior in terms of their capacity to mediate cytotoxic functions upon direct and anti-HIV-1 antibody dependent stimulation39. We studied NK cell functionality in both a direct manner (K562 co-culture) and an indirect manner (ADCC). Our results showed a trend towards increased direct and indirect killing of target cells at Vac#2 + 1w. However, this effect did not remain after ATI, which is in line with the observation that PLWH with high viremia show decreased cytotoxicity via both direct and indirect pathways37.
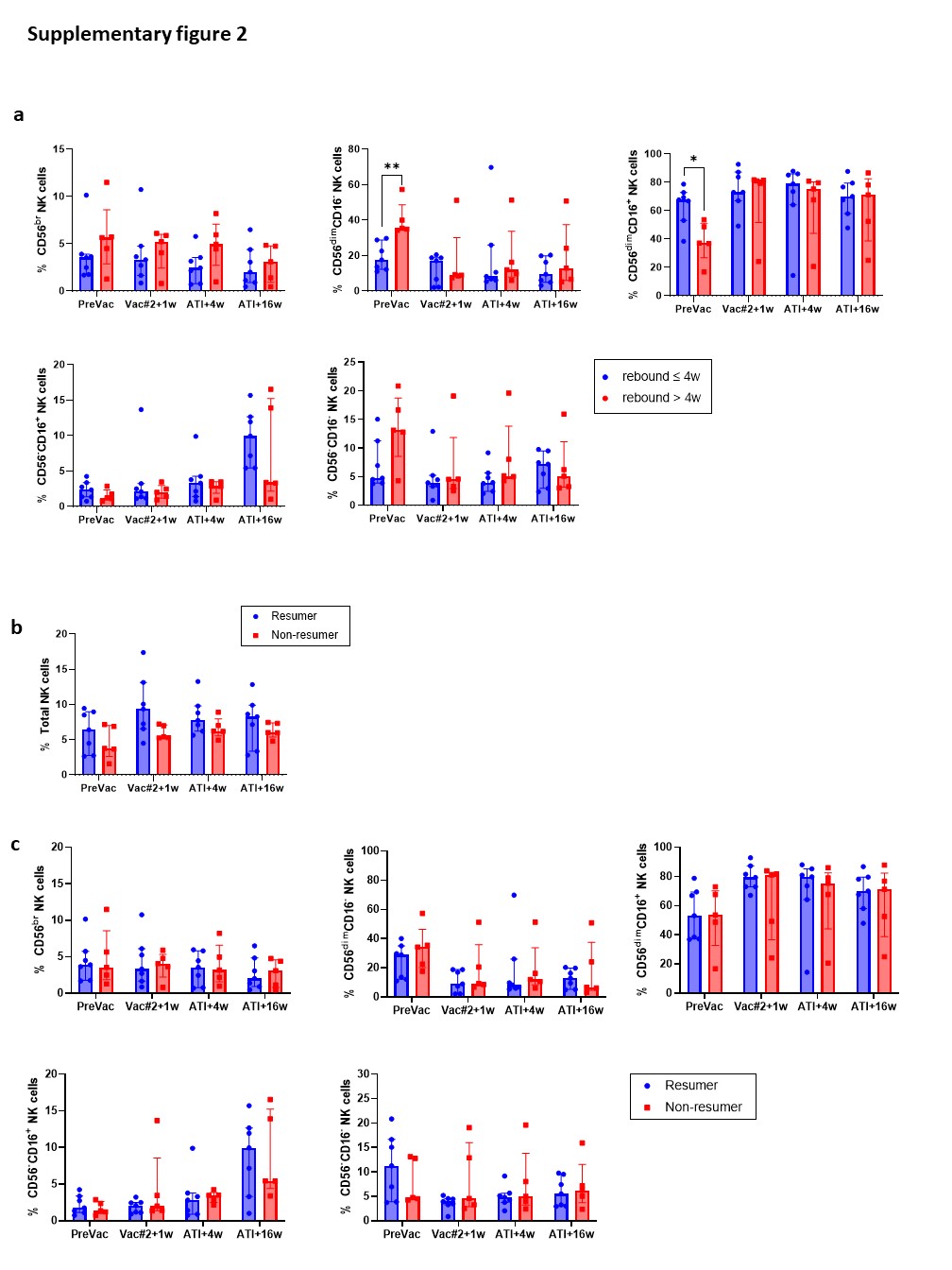
In the context of cancer immunotherapy, clinical trials suggest that DC-based vaccination results in increased lytic activity and profound effects on NK cell phenotype, which correlate with clinical responses40–43. Although we previously observed vaccine-induced antigen-specific CD8+ T-cell responses on the one hand6, and profound changes in phenotype and functionality of NK cells upon vaccination, on the other hand, no prolonged time off ART following ATI was observed compared to historical controls. Importantly, the trial was performed in 2007 meaning that guidelines to restart ART following ATI were less strict compared to current criteria. The timing to restart ART was based on the judgement of the treating physician, which could introduce a level of bias. Viral rebound occurred 4 weeks (median) after ATI, whereas ART was restarted many weeks afterwards (28–352 weeks after ATI). Nevertheless, NK cell frequency, phenotype and functionality did not seem to be predictive for time to viral rebound. Another important limitation of this study is the lack of a time point after the fourth vaccination where all participants are still virally suppressed.
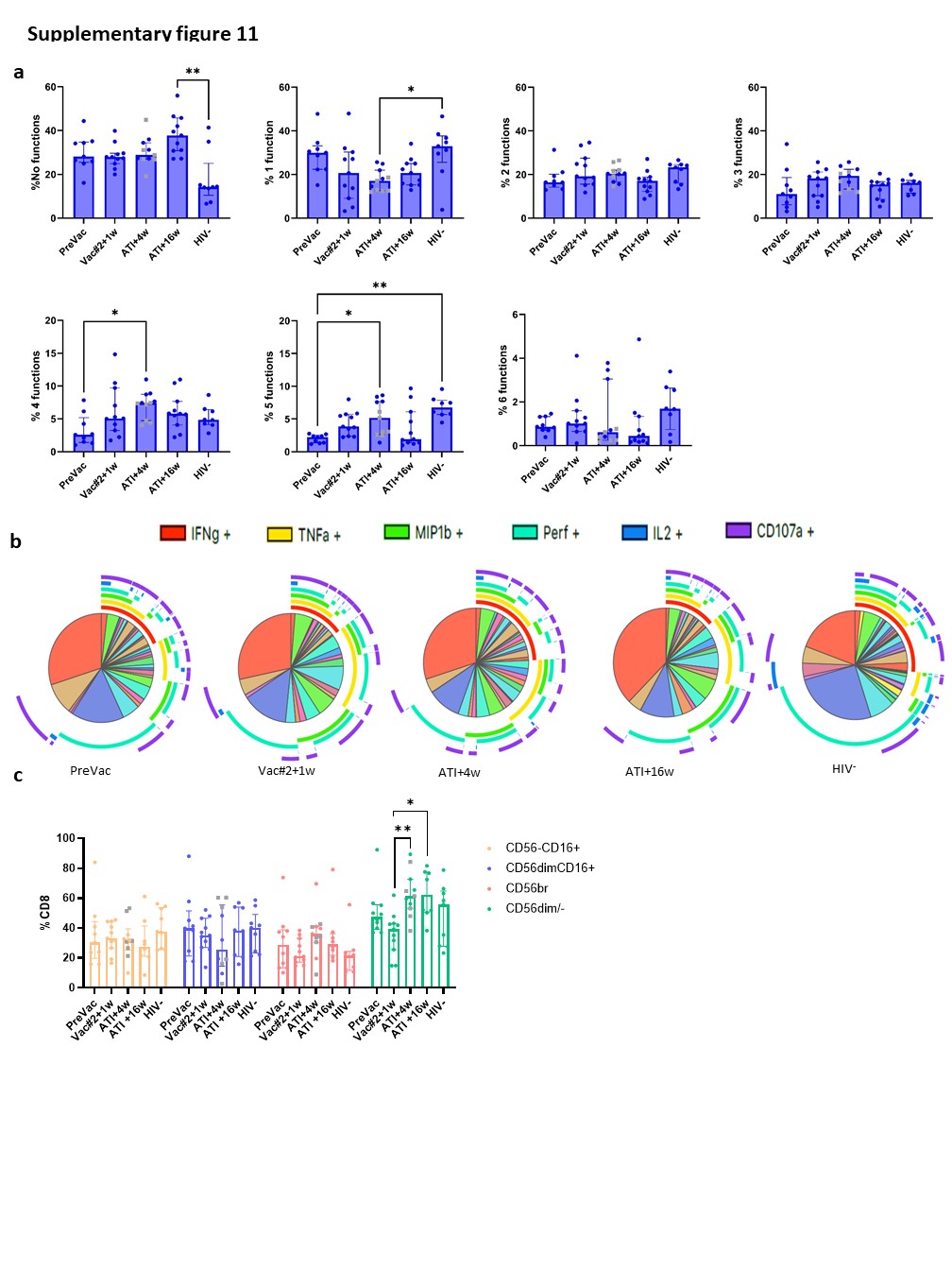
T-cell polyfunctionality is a well-established concept within the field of cancer and HIV-1 immunology. However, for NK cells, this concept has not been extensively studied and the few studies that have addressed “polyfunctionality” generally limit their analysis to a restricted number of markers. Thus, we are the first to perform a bona fide polyfunctionality analysis on NK cells, based on the extended analysis of T-cell polyfunctionality published by Hersperger et al. 44. We found that, at Vac#2 + 1w, mainly those NK cells with higher functionality (4–5 functions) were significantly increased in number, which in T cells has been shown to correlate with better control of viral replication and slower disease progression. Interestingly, Ahmad et al. showed that CD8+ NK cells exhibit higher polyfunctionality, which was also associated with a slower disease progression in untreated PLWH30. At Vac#2 + 1w, we found a positive correlation between frequency and polyfunctionality of CD8+ cytotoxic NK cells which was absent at baseline.
In order to eliminate HIV-infected cells, NK cells need to be functional. ICMs are an important class of receptors hampering this functionality upon interaction with the respective ligand. In contrast to CTLs, in NK cells, the expression of ICM and the phenomenon of exhaustion have not been studied in great detail, neither in the context of cancer nor in PLWH. Norris et al. reported that the frequency of PD-1+ NK cells increases during HIV-1 infection and is only partially restored upon ART initiation resulting in reduced proliferative and functional capacity45. At Vac#2 + 1w, we observed a significant decrease in ICM which did not persist after ATI. Immune checkpoint inhibitors are rapidly becoming the treatment of choice for several cancer types, whereas only few clinical studies have been performed in the context of HIV-1. Recently, it was shown that anti-PD-1 treatment also has latency reversal effect46. Combining latency reversal with DC based vaccination is an interesting approach worth exploring in the context of HIV cure. In this regard, Gay et al. recently showed that Vorinostat combined with an autologous DC-based vaccine (AGS-004) resulted in increased T-cell responses and a reduction in viral reservoir size5,47. However, NK-cell responses were not investigated in this study.
NK cells are normally excluded from LNF, so in order to control the viral reservoir NK cells need to migrate towards LNF. Expression of CD62L facilitates homing to secondary lymphoid organs by initiating the tethering and rolling on the endothelium. Martin-Fontecha et al. showed that injection of mature DCs induced migration of NK cells to LNF in a CD62L dependent manner, which ultimately leads to Th1 polarization48. Although CD62L has been suggested to be expressed mainly on CD56dim NK cells, we found the highest expression on the CD56br subset. While the percentage of CD62L and CCR7 increased on cytotoxic NK cells at Vac#2 + 1w, CXCR5 expression was not altered. This could be attributed to the fact that we studied peripheral blood NK cells instead of lymphoid NK cells. It has been shown that a significant number of blood NK cells in African Green monkeys is strongly positive for the LNF homing marker CXCR5, leading to control of the viral reservoir49. Similar data were recently obtained in a model for chronic SHIV infection. Moreover, in human samples, significantly increased numbers of CXCR5+ NK cells were found in LNF from PLWH compared to healthy donors50,51. The size of the viral reservoir was, however, not determined in this study.
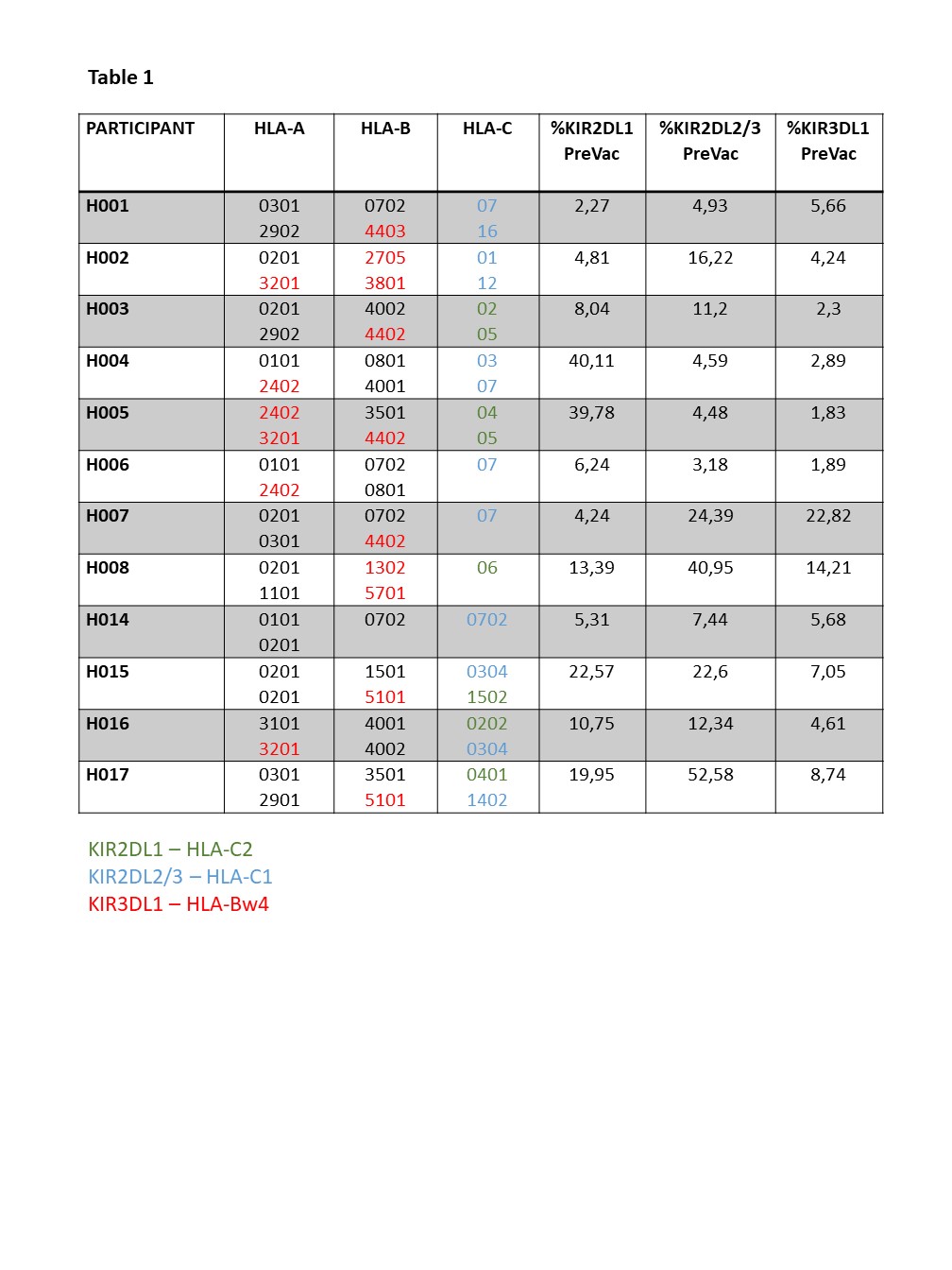
Immunological memory has traditionally been known to be an “exclusive” hallmark of adaptive T and B cells. However, this paradigm has been challenged by increasing evidence indicating that NK cells have certain features of adaptive immunity as well. Different groups have identified adaptive NK cells in mouse models for hapten-induced hypersensitivity and viral infections (MCMV, HIV-1 and Influenza)52–54. In a primate model of SIV, these adaptive NK cells were shown to lyse Gag- and Env-pulsed autologous DCs up to 5 years after vaccination28. In humans and primates, NKG2C is thought to be involved in the antigen specificity (presented by HLA-E)55. However, this needs to be further clarified. Our results showed no significant increase in memory-like NK cells, but a trend towards a positive correlation with direct and indirect cytotoxicity was observed. Importantly, the CMV status of participants should be taken into account as this might trigger memory-like NK cell responses56. Unfortunately, this information was not available for the vast majority of our study participants. Moreover, peripheral blood might not be an ideal site to study adaptive NK cells as these cells have been shown to reside in the liver and spleen in SIV models28.
The administration route and mode of vaccine-mediated antigen presentation are critical for DC stimulation and subsequent crosstalk with NK cells. In the present study, an ex vivo loaded autologous DC vaccine was administered both subcutaneously and intradermally, whereas peptide-loaded DC administered via subcutaneous injection is believed to induce the best NK cell responses in mice48,57. In addition, DCs that have been pulsed with the HIV-1 accessory protein Nef have previously been shown to increase IFN-γ production by CD56bright NK cells as well as reduce cytotoxicity of CD56dimCD16+ NK cells in vitro58.
Ex vivo loading of autologous DCs with HIV-1 antigens is quite costly and requires extensive expertise and infrastructure. This has led to the development of nanoparticle based mRNA vaccines, which have been proven to be very effective in the context of SARS-CoV-259. Moreover, an increase in NK cell frequency with increased IFN-γ production was observed following BNT162b2 vaccination, which suggests it is worth to investigate these nanoparticle based mRNA vaccines against HIV-1 as well60.
In conclusion, our results demonstrate that DC-based immunotherapy resulted in an increase in the frequency of cytotoxic NK cells. Moreover, NK cells had a more licensed and migratory profile which translated into increased cytotoxicity and polyfunctionality. Since no delay in time to viral rebound was observed, these results warrant further research to optimize the efficacy of vaccine-induced NK-cell responses. Overall, this work highlights the importance of in-depth analysis of NK-cell frequency, phenotype and functionality after and/or during DC-based immunotherapy, which has an impact on future HIV-1 cure research. Currently, the main focus of DC-based immunotherapy lies on the improvement of adaptive immune responses and our results can extend this to NK-cell mediated responses. This is not only of great importance in the field of infectious diseases but can be easily expanded to other fields, such as cancer immunotherapy, as well.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}