Genome-wide identification of VvHsp20 gene family in grape
A total of 61 VvHsp20 genes were obtained by Hidden Markov Model (HMM) analysis. The presence of an ACD domain was confirmed by submitting the protein sequences to CDD, Pfam, and SMART database. The sequences without the typical ACD domain were discarded. A total of 48 sequences were retained and confirmed as grape Hsp20 after removing the sequences with a molecular weight beyond the 15–42 kDa. Detailed information on physicochemical properties of these Hsp20s are listed in Table 1. The length of the VvHSP20 proteins varied from 136 (VvHSP20-47 and VvHSP20-48) to 365 amino acids (VvHSP20-41); the molecular weights of VvHSP20s were from 15.27 kDa (VvHSP20-30) to 40.59 kDa (VvHSP20-41). The predicted pI values of VvHsp20s ranged from 4.68 (VvHsp20-41) to 9.48 (VvHsp20-20).
Phylogenetic analysis of VvHsp20 genes
An unrooted Neighbor-Joining (NJ) phylogenetic tree was constructed based on the alignment of amino acid sequences of Hsp20 from grape, Arabidopsis, tomato (Fig. 1). In total, 19 sequences from Arabidopsis, 26 sequences from tomato, and 48 sequences from grape were assessed in the phylogenetic tree. according to the phylogenetic and the subcellular localization analysis, the grape Hsp20 protein are divided into 11 subfamilies (CI, CII, CIII, CV, CVI, CVII, MI, MII, ER, CP, and PX/Po) (fig. 1, Table 1). Clustering of the subfamilies in grape is largely consistent with the subcellular localization, i.e., the proteins in the same cluster were located in the same subcellular sites. Specifically, six Hsp20 subfamilies (CI-CVI), MTI and MTII subfamilies, CP, ER and PX /Po localize to the cytoplasm/nucleus, mitochondria, chloroplast, endoplasmic reticulum and peroxisome, respectively. The 93 Hsp20s were classified into 14 distinct subfamilies, except for the unclassified VvHsp20s (VvHsp20-15, VvHsp20-16, VvHsp20-38, and VvHsp20-41), the subcellular localization of which could not be predicted using the online tool Protcomp. Most of the VvHsp20s, including 33 out of 44, were classified into CI–CVII, which suggested that the cytosol may be the primary functional site of plant Hsp20s.
Characterization of the amino acid sequences and gene structure of VvHsp20s
As shown in Fig. 2a, 48 VvHSP20s were divided into 11 subgroups, except for the unclassified Hsp20 (VvHsp20-15, VvHsp20-16, VvHsp20-38 and VvHsp20-41). Ten conserved motifs of VvHsp20 proteins were identified by the MEME website and listed in Table 2. The lengths of these conserved motifs ranged from 6 to 60 amino acids (Fig. 2b, table 2). ACD consists of two conserved regions, CRI of β2, β3 and β4, and CRI of β7, β8 and β9, separated by a variable length hydrophilic region β6 loop (Fig. 3). VvHsp20-2, 3, 39, 40, 47 and 48 lacked the β6-loop. VvHsp20-36 lacked the β-strands 4. The different components of the ACD domain suggest functional diversity among VvHsp20s. The same group of VvHsp20 proteins in the phylogenetic tree had the same motif, which indicated that they were highly conserved.
Next, we analyzed gene structure in order to better understand hsp20s. Among the VvHsp20s genes, 24 (50.0%) were intronless, and 21 genes (43.8%) possessed one intron. VvHsp20-38 (2 introns), VvHsp20-34 (3 introns), and VvHsp20-20 (5 introns) had two or more introns (Fig. 2c). Genes of the same subgroup had the same intron phase, which indicated that the structure was quite conserved over evolution.
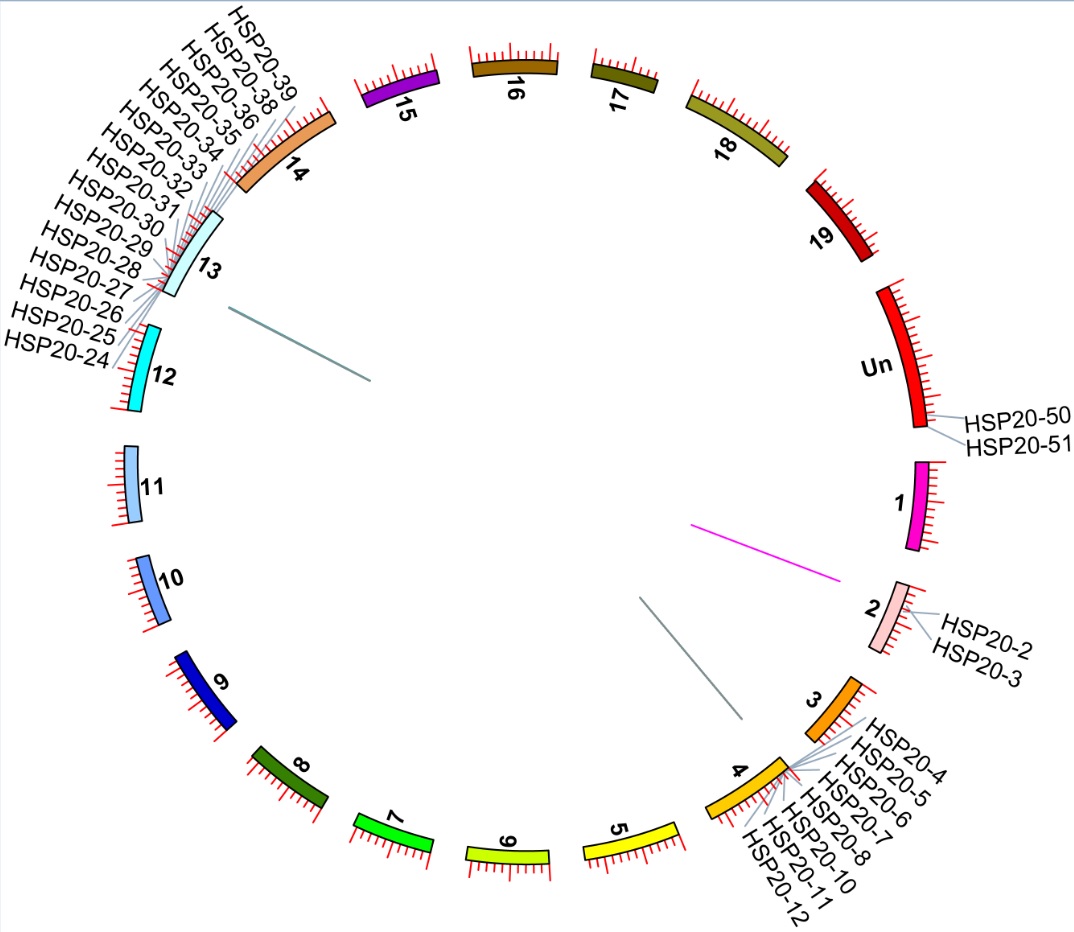
Chromosomal location and gene duplication of VvHsp20
The 48 VvHsp20 genes were distributed on 12 grape chromosomes (Fig. 4). most of the VvHsp20 genes were present on chromosome 4 (10 genes) and chromosome 13 (16 genes), while each of the remaining 10 chromosomes had one or two genes. Both tandem and segmental duplication contribute to the production of gene families during the process of evolution. Thus, potential duplication events of VvHsp20 genes were analyzed. In total, four groups of VvHsp20 genes (VvHsp20-2, 3; VvHsp20-4, 5, 6, 7, 8, 9, 10, 11; VvHsp20-23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 35, 36 and VvHsp20-47, 48) were identified as tandem duplication genes (Additional file 1: Figure S1). Furthermore, none of the genes were suggested to be products of segmental duplication. Based on the above results, we inferred that tandem duplication played an important role in the expansion of the VvHsp20 family in grape.
Analysis of cis-element in VvHsp20 gene promoters
To understand the possible role of cis-regulatory elements of VvHsp20, the promoter sequences (comprising −2000 bp upstream of the translation start site) of 48 VvHsp20 genes were submitted into PlantCARE to detect the cis-elements. three categories of cis-elements, including phytohormone responsive, abiotic and biotic stress-responsive, and plant development-related cis-elements were identified and are shown in Fig. 5. Among the three categories of cis-elements, the phytohormone responsive category accounts for the highest proportion. In this category, cis-acting elements were widely present in the promoter region, including auxin responsive (TGA-element and AuxRR-core), gibberellin-responsive elements (GARE-motif, P-box, and TATC-box), ethylene-responsive (ERE), MeJA-responsive (TGACG-motif and CGTCA-motif), abscisic acid-responsive (ABRE), and salicylic acid-responsive (TCA-element). Among these elements, ABRE and ERE accounted for the largest part of the phytohormone responsive category. In the abiotic and biotic stress-responsive category, stress response-related cis-elements, such as HSE1 (heat stress), WUN motif (wound-responsive element), TC-rich repeats (stress response), LTR (low temperature-responsive), ARE (anaerobic induction), and GC-motif (anoxia) were detected. In the last category, plant development-related elements, including meristem expression (CCGTCC-box and CAT-box), circadian, zein metabolism regulation (O2-site), cell cycle regulation (MSA-like), differentiation of the palisade mesophyll cells (HD-Zip 1), and endosperm expression (AACA_motif and GCN4_motif) were identified. In addition, most of the VvHsp20 genes possessed W boxes and MYB binding sites, including CCAAT-boxes.
Expression patterns of VvHsp20s in response to H2O2 treatment
There is a close relationship between gene expression and function. To determine the functions of VvHsp20s in grape, a heatmap of 47 VvHsp20 genes was constructed using FPKM values from RNA-Seq data in control and H2O2-treated berries of ‘Kyoho’ (Fig. 6, sampling period is described in Materials and Methods and table 3). The expression level of hsp20-33 was extremely low and not detected by RNA-Seq analysis during fruit development. Most VvHsp20s were down-regulated after treatment, especially at the fourth period. However, the opposite trend was also observed for a few genes, including hsp20-13, hsp20-20, and hsp20-30. These results indicated that most of the VvHsp20 genes responded to H2O2 treatment, and the response mechanisms of different VvHsp20 genes to H2O2 were different.
based on the statistical significance of the gene expression levels from the RNA-Seq analysis and the partitioning of the clusters of genes from the phylogenetic analysis, 14 differentially expressed VvHsp20 genes were selected to be further validated by qRT-PCR in response to control and H2O2 treatment (Fig. 7). Consistent with the RNA-Seq data, the expression level of most genes decreased after the treatment. Besides hsp20-31, the relative expression levels of the remaining 13 genes were extremely down-regulated at the fourth period. It is worth noting that VvHsp20-17 and VvHsp20-25 were hardly expressed after treatment. Similar expression patterns were revealed within the tandem duplicated gene groups (VvHsp20-25 and VvHSP20-28). The similar expression patterns indicated that the tandem duplicated VvHSP20 genes had similar functions and structures. Members of the CI subgroup (VvHsp20-24, VvHsp20-25, VvHSP20-28, and VvHsp20-31) had similar expression patterns after the treatment, which suggested that they had similar functions in response to H2O2 treatment.
Expression patterns of ABA-related genes in response to H2O2 treatment
It is well known that ABA plays an important role in grape [23, 24]. In the previous study [1], H2O2 treatment was shown to promote the early fruit ripening of ‘Kyoho’. To further explore the role of ABA in this process, RNA-Seq and qRT-PCR were performed to examine the expression analysis of ABA-related genes. As shown in Figure 8, the expression patterns of the ABA synthesis-related gene (NCED3) and degradation-related gene (CYP707A4) were different following H2O2 treatment. Compared with the control, the expression level of NCED3 reached the highest level at veraison (H3 stage), then decreased at the H4 stage. On the contrary, the expression level of the CYP707A4 gene increased rapidly after treatment and reached its lowest level at veraison. The changes in the expression levels of ABA-related genes indicated that H2O2 may regulate fruit development possibly through control of ABA catabolism and biosynthesis.
{kind=link}