K 2 SO 4 did not impact the mortality and infectivity of M. graminicola at low concentrations
The direct toxicity of K2SO4 to M. graminicola was determined by calculating the mortality of nematodes at different concentrations of K2SO4 solution 72 h after treatment. Although the mortality of nematodes increased along with the increase of K2SO4 concentrations ranging from 0.125 mM to 8 mM, there was no significant difference compared to the control water treatment (5.0 ± 0.7%), especially at 0.125 mM K2SO4 concentration (5.7 ± 1.4%) (Fig. 1a). These results indicated that the K2SO4 was not toxic to M. graminicola at low concentrations within 72 h.
To determine whether K2SO4 directly impact the infection and development of M. graminicola, the newly hatched second stage juveniles (J2s) were soaked in 0.5 mM K2SO4 solution for 48 h before inoculation. At 14 days post inoculation (14 dpi), no difference was observed in the total numbers of juveniles and adult females between the K2SO4-soaked and the water-soaked groups, which indicated that there was no direct effect of K2SO4 on the infection of M. graminicola. At the same time, most nematodes developed into adult females, and there was no significant difference in the numbers of third stage juvenile (J3), fourth stage juvenile (J4) or adult females between the two groups (Fig. 1b, c). These results indicated that K2SO4 did not suppress the development of M. graminicola.
K 2 SO 4 treatment induced resistance of rice to M. graminicola
To evaluate the resistance of rice against M. graminicola induced by K2SO4,different concentrations of K2SO4 solution treated the rice roots 1 d before inoculation, and preliminary plant infection rate was assayed at 14 dpi. The results showed that compared to the water treatment, K2SO4 treatment could effectively reduce the numbers of both galls and nematodes (juveniles and adult females) at the concentration of 0.125 mM, and there was no significant difference among the K2SO4 treatments. After applying with K2SO4 solution at a concentration of 0.5 mM, the numbers of both root galls and nematodes were decreased significantly (57.2 ± 4.4%, 59.2 ± 6.6%) at 14 dpi (Fig. 2a, b). In addition, compared to the untreated control plants, the development of nematodes treated with K2SO4 was evidently delayed. At 14 dpi, the proportion of adult female in plants treated with K2SO4 (70.9 ± 5.6%) was significantly lower than that of plants in water control (90.7 ± 5.1%), and the proportion of juveniles (J3 + J4) (27.0 ± 6.3%) in plants treated with K2SO4 was higher than that of control plants (6.0 ± 3.2%) (Fig. 2c, e). These results indicated that the low concentrations of K2SO4 could effectively reduce the infection of nematodes and delay the development of nematodes. Therefore, the 0.5 mM K2SO4 solution was used in all the subsequent experiments. We also calculated the male/female ratio of nematodes, indicating that the ratio in rice roots treated with K2SO4 was lower than that in the untreated roots (Fig. 2d). Meanwhile, there was no significant difference in the fresh weight and height of rice shoots or roots between K2SO4 treatment and water treatment (Fig. 2f, g), suggesting that application of K2SO4 had no toxic effect on rice growth. All these results indicate that the application of K2SO4 can induce rice resistance against M. graminicola.
K 2 SO 4 -induced resistance of rice to M. graminicola was achieved by neither impacting the attractiveness of rice roots to nematodes nor inhibiting the developmental phenotype of giant cells
In nature, attraction of the host roots to nematodes is of great importance in establishing infection [13]. Here, the nematodes in the range of 5 mm around the root tips treated with K2SO4 solution were counted. At 10 hours post inoculation (hpi), the numbers of nematodes attracted to the K2SO4 treated tips (26.2 ± 3.1) were not different from the water treated tips (25.7 ± 2.8) (Fig. 3a, b). The results indicated that K2SO4 treatment did not affect the attraction to M. graminicola.
The invaded J2 can induce the plant to form giant cell at the feeding site by secreting effectors from esophageal gland through stylet [14], and to acquire the nutritional needs of subsequent growth and development. Then the giant cells develop to form the root galls. In this experiment, the root galls of 7 dpi were collected and used for histological observation. The average cross-sectional area of giant cells in K2SO4 treated group was not significantly different from the untreated group (Fig. 3c, e). We also counted the numbers of giant cells in feeding sites; there was no significant difference between the two treatments (Fig. 3d). All these results indicated that the application of K2SO4 did not influence the development of giant cells.
K 2 SO 4 induced resistance to M. graminicola in Rice by priming basal defense
Callose deposition is one of the markers of plant defense response, which can play a role in resistance to pathogen infection [15]. We observed the presence of callose in the gall tissue induced by RKNs after staining with aniline blue. The results showed that compared with the water treatment group, more callose was deposited in the K2SO4 treatment group, and the callose spots were larger and more dense (Fig. 4a, b), with 67.9% of increase in the average area of callose spots (Fig. 4a). These results suggest that increased callose deposition is one of the pathways of K2SO4-induced resistance in rice plants.
Hydrogen peroxide (H2O2) is a signaling molecule that mediates plant defense response to biological or abiotic adversity [16]. Our analyses indicated that the H2O2 response of rice roots could also be induced by K2SO4 alone, because the H2O2 level at 24 hpi was significantly higher than that of the control group. In the case of nematode challenge, compared with the rice without K2SO4 treatment, the level of H2O2 in treated rice was increased by 78.2% and 118.7% at 8 and 24 hpi, respectively, but it decreased at 72 hpi (Fig. 4c). In addition, the expression of OsRbohB, an H2O2 synthetic gene known to be involved in plant immune response [17], was increased in the K2SO4-treated rice at 8 and 24 hpi. Especially, at 8 hpi, the expression level of OsRbohB in the inoculated plant treated with K2SO4 was significantly higher than that in the non-treated (P ≤ 0.05) (Fig. 4d), which was consistent with the results of H2O2 content (Fig. 4c). These results suggest that the application of K2SO4 can induce defense against M. graminicola by activating the rapid production of reactive oxygen species (ROS).
K 2 SO 4 treatment promoted the expression of disease resistance-related genes in inducing rice resistance against M. graminicola
While the plant is attacked by the pathogens, it will activate the autoimmune response system to defend. Immune response is a complex process involving the transcription and expression of many disease resistance-related genes [18]. We analyzed the expression of several common disease resistance-related genes in different treatments and at different time points.
Firstly, we analyzed two salicylic acid (SA) pathway-related genes: SA transcription factor OsWRKY45 and biosynthesis gene OsICS1. The expression of both OsWRKY45 and OsICS1 was significantly increased by K2SO4 treatment alone at 72 hpi, however, after inoculation, K2SO4 treatment only induced OsWRKY45 at 24 hpi. OsICS1 was not prominent at any time points (Fig. 5a). Therefore, SA pathway is not the main pathway in K2SO4-induced rice resistance against M. graminicola. Secondly, we analyzed two jasmonic acid (JA) pathway-related genes and two ethylene (ET) pathway-related genes. After treatment with K2SO4, JA transcription factor gene OsJAMYB was significantly increased in both inoculated and non-inoculated groups at 8 hpi (Fig. 5b). The expression pattern of JA biosynthesis gene OsAOS2 was similar to that of OsJAMYB, but it was induced more than OsJAMYB, at 8 hpi (Fig. 5b). Although the ET signaling gene OsEIN2 was not expressed much, it was expressed significantly higher in K2SO4-treated inoculated rice than in untreated. The expression of OsEIN2 was slowly increased at all the three time-points, while the expression of ET biosynthesis gene OsACS1 was increased sharply in the K2SO4-treated and MG-inoculated group at 72 hpi, which was 6.98 times higher than that of untreated but MG-inoculated group (Fig. 5c). These results suggest that K2SO4 plays an active role in inducing the expressions of JA and ET pathways-related genes in rice resistance to RKNs. Thirdly, we analyzed the expression of pathogenesis-related (PR) genes. The expression of both OsPR1a and OsPR1b in the K2SO4-treated rice was significantly increased compared to the untreated group at 8 hpi. The expression of OsPR1b continued to be increased in the inoculated rice treated with K2SO4 after 8 hpi, and was still significantly higher than that in the untreated at 24 hpi (Fig. 5d). These results suggest that K2SO4 positively regulates the expression of these two genes in rice resistance to RKNs.
Finally, we analyzed the expression of two brassinolide (BR) pathway-related genes. The expression of BR receptor gene OsBRI1 was significantly increased in both inoculated and uninoculated K2SO4-treated groups at 24 hpi and 72 hpi, while the expression level of BR biosynthesis gene OsDwarf in the inoculated group treated with K2SO4 was significantly higher than the untreated at 24 hpi (Fig. 5e). This indicates that BR pathway also plays an active role in the resistance process, and K2SO4 positively regulates the expression of BR pathway-related genes.
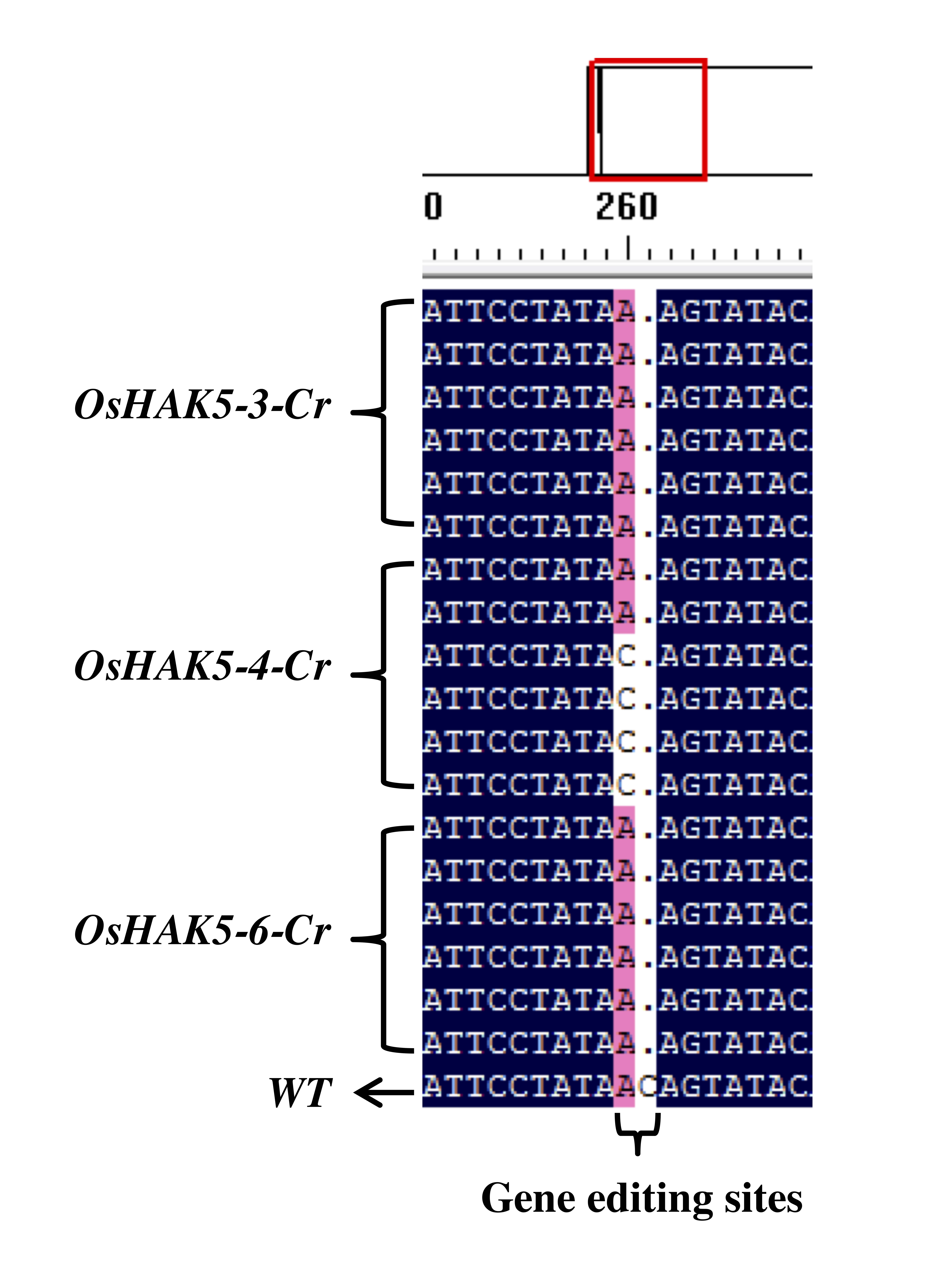
The resistance of rice against nematodes induced by K 2 SO 4 required the participation of potassium absorption and transportation systems
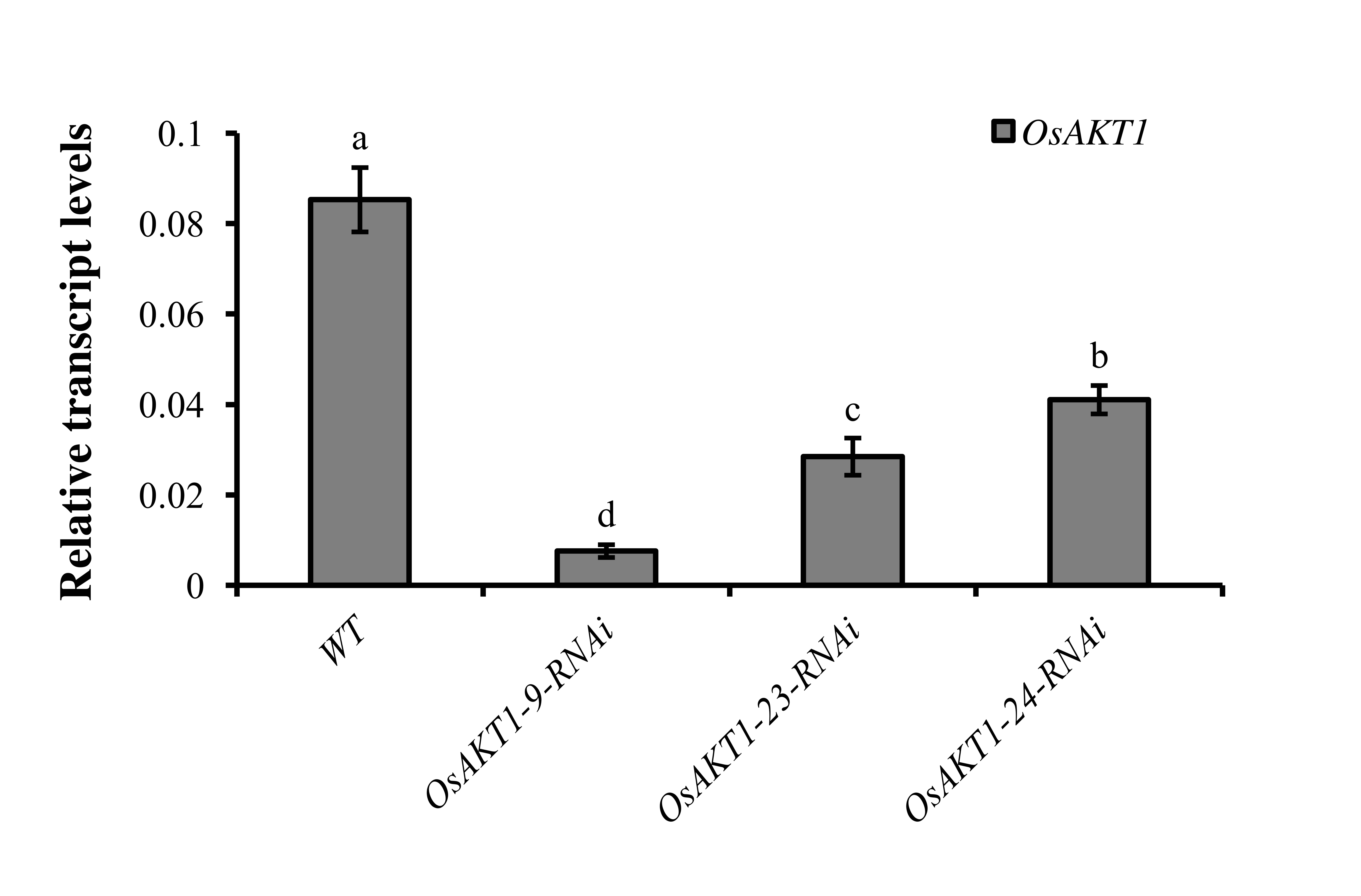
To determine the role of potassium (K+) absorption and transportation systems in disease resistance process, we first compared the expression of K+ channel gene OsAKT1 and K+ transporter gene OsHAK5 between K2SO4 application and non-application, and between nematode inoculation and non-inoculation. The expression of OsAKT1 in the inoculated rice treated with K2SO4 was higher than that in both the untreated and in the non-inoculated within 72 h, especially at 8 h and 72 h (Fig. 6a). This suggests that OsAKT1 expression is affected by K2SO4 and nematodes, which may be involved in the resistance process of rice against nematodes induced by K2SO4. Meanwhile, the expression of OsHAK5 was up-regulated in all the treatments within 24 h after inoculation. Although there was no significant difference among the treatments, the expression of OsHAK5 was slightly higher in the inoculated rice than in non-inoculated, and the expression of OsHAK5 was slightly higher in the inoculated rice treated with K2SO4 than in the non-treated (Fig. 6a). This indicates that nematode infection and K2SO4 treatment have slight effects on OsHAK5.
Subsequently, we inoculated nematodes on the rice lines deficient in these two genes. Interestingly, in the presence of K2SO4 at a concentration of 0.5 mM, both of the two defective plants (OsAKT1-RNAi and OsHAK5-Cr) became more susceptible than the wild-type plants. Specifically, compared with the two defective plants, the numbers of both galls and nematodes (juveniles and adult females) in the wild-type plants roots were reduced, and the proportion of juveniles (J3 + J4) were increased, while the proportion of adult females was reduced. (Fig. 6b, c). These results indicate that these two genes are associated with the inhibition of the infection and development of nematodes in rice. On the hand, in the case of K+ starvation, the susceptibility of the two defect types was still higher than that of wild type (Fig. 6b, c). These results suggest that these two genes also play an independent role in nematode resistance. However, without K2SO4 treatment, the relative susceptibility of OsAKT1 and OsHAK5 deficient lines was weaker than that with the K2SO4 treatment (Fig. 6b, c). These results suggest that although both OsAKT1 and OsHAK5 have independent roles in the process of disease resistance, both were involved in K2SO4-induced resistance of rice against M. graminicola.
{kind=link}
{kind=link}