MEF2D participated in the inflammatory response of BV2 cells
Previous studies showed MEF2D participated in MPTP-induced microglial activity. This prompted us to explore the transcriptional regulation network of MEF2D. We chose the model of LPS-induced BV2 microglia cell line, a widely used model of inflammation. After LPS treatment, the level of MEF2D was gradually increased, and reaching a significantly higher level at 24 h (Figure 1A left). As MEF2D is a transcript factor, the nuclear and cytoplasm were separated to confirm the nuclear distribution of MEF2D (Figure 1A right). The elevated MEF2D after LPS treatment was contributed only in nuclear, while the cytoplasmic level didn’t change, indicating that MEF2D may play a transcript role in inflammation.
To verify the regulating effect of MEF2D in inflammation, we used lentivirus vector to get stable BV2 cells of shMEF2D and OE-MEF2D. After LPS stimulation, the expressions of proinflammatory factors NLRP3 and iNOS were significantly promoted by MEF2D knockdown, and diminished by overexpression of MEF2D compared with NC (Figure 1B). As IL-4 could inhibit macrophage production of proinflammatory cytokines, we also administered IL-4 and found that the MEF2D level did not change significantly compared to untreated groups in different stable cell lines (Figure 1B). These data indicate that MEF2D is more inclined to participate in the proinflammatory reaction process.
Morphology is also an important indicator of the activation state of microglia. In the resting state, BV2 microglia cells were round with small cell bodies, while under LPS stimulation, the morphology changed significantly: the cell body became larger and the protrusions became more and thicker. It is interesting to note that knockdown MEF2D made BV2 cells turbulent on the baseline. The rate of activated shMEF2D BV2 cells was significantly higher than NC at 12 h after LPS treatment. On the other hand, overexpression of MEF2D mitigated the activation state even under LPS treatment (Figure 1C). Immune phagocytosis is another main feature of activated microglia. The data showed consistent results that knockdown MEF2D enhanced immune phagocytic function of BV2 cells, while overexpression of MEF2D alleviated the immunophagocytic activity to certain extent (Figure 1D). These data suggest that MEF2D might act an inhibitor of excessive inflammation to be tightly involved in the regulation of inflammatory homeostasis.
MEF2D knockdown promoted pro-inflammatory response of BV2 cells
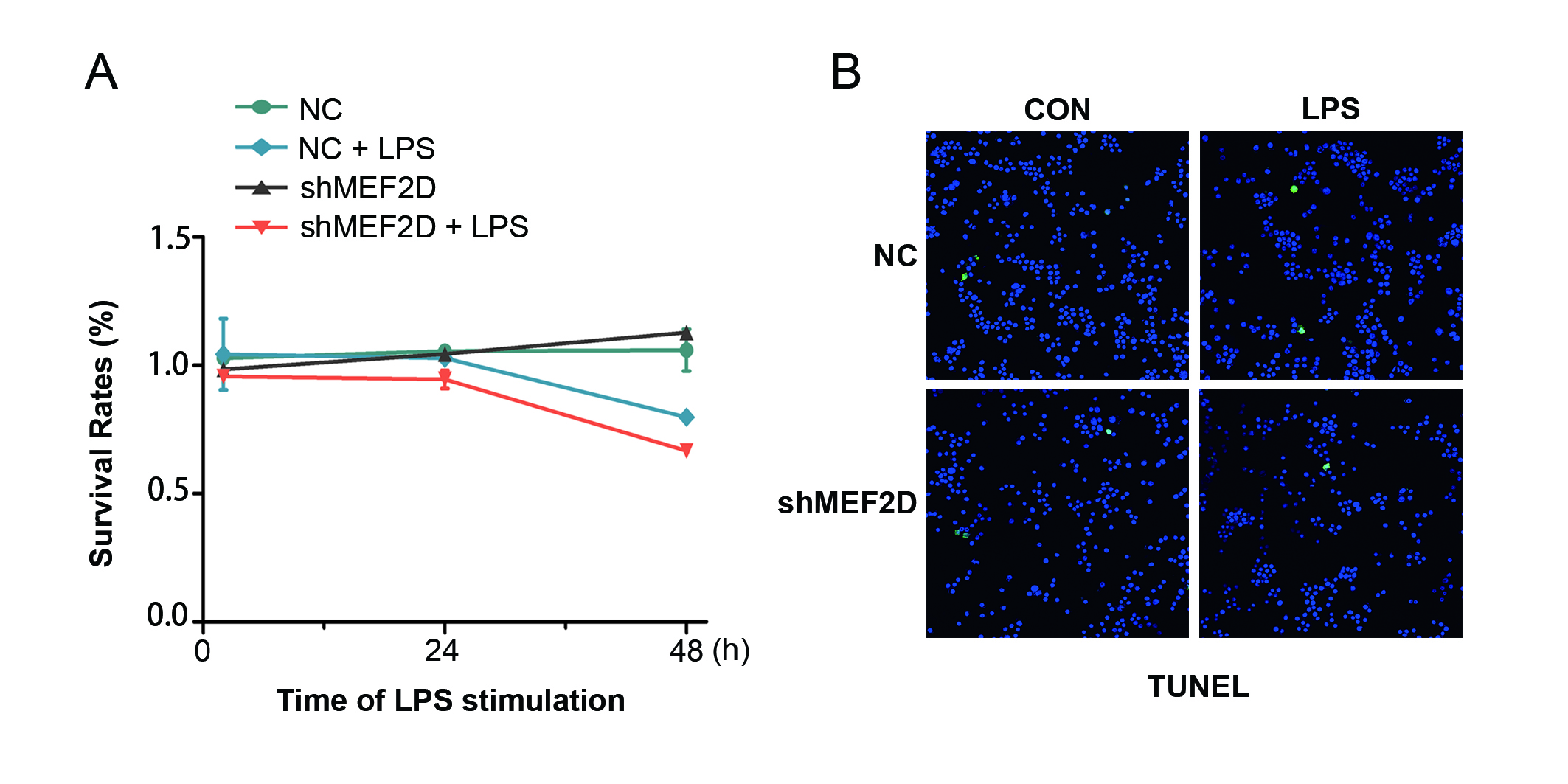
Our above data showed that MEF2D may participate in the inflammatory response of microglia. To further test this proposal, we used steady shMEF2D BV2 cells, of which the knockdown efficiency was about 80% of the background compared to NC. Even with LPS stimulation for 24 h, the expression of MEF2D in shMEF2D BV2 cells was significantly lower (Figure 2B and E). The MTT and TUNEL assays were performed, and the data showed that knockdown MEF2D didn’t affect the survival and proliferation of BV2 cells, even under stress condition of LPS (Figure S1A and B).
To role out the exact influences of MEF2D in activated BV2 cells, we performed time course studies of several key inducible inflammatory factors in shMEF2D BV2 cells under LPS treatment. NLRP3 is an innate immune pro-inflammatory factor to mediate the cleave-mature process of IL-1β. Its function is crucial for the regulation of neuroinflammation mediated by microglia. Knockdown MEF2D significantly promoted the expression of NLRP3 after LPS treatment for 8 h, and the trend lasted for at least to 24 h in BV2 cells (Figure 2A and B). The secretion level of downstream cytokine IL-1β of shMEF2D was significantly higher after 12 h of LPS stimulation compared to NC (Figure 2C). The mRNA and protein levels of another pro-inflammatory factor iNOS were also increased markedly by knockdown MEF2D compared with NC after 8 h treatment with LPS (Figure 2D and E). On the contrary, the protein but not the mRNA level of the anti-inflammatory factor nuclear factor-erythroid 2-p45 derived factor 2 (Nrf2) was deceased (Figure 2D and E). These results strongly support the fact that knockdown MEF2D markedly enhances the susceptibility and the pro-inflammatory response of BV2 microglia cells.
Differently expressed genes in response to MEF2D knockdown enriched in the immune system in BV2 cells
To investigate the regulatory function of MEF2D in microglia, we used RNA-seq and analyzed the global gene expression patterns in LPS-stimulated BV2 cells with or without steady MEF2D knockdown. To monitor the changes of inflammatory factors in the acute stage, we chose the 12 h after LPS treatment to perform transcriptome sequencing. The knockdown efficiency and the transcriptional activity were tested for the quality control (Figure 3A and B). The RNA-seq transcriptional analysis was performed using three independent samples (biological replicates) of each treatment: NC, NC + LPS stimulation, shMEF2D, shMEF2D + LPS stimulation. The principal components analysis (PCA) showed a good separation and a high level of consistency between biological replicates of the same population in BV2 cells (Figure 3C). Using a false discovery rate, P≤0.01, and fold change≥1.5 log2 as the cutoff values, we identified 310 differently expressed genes in knockdown MEF2D cells relative to controls without LPS treatment (Figure 3E). The genes were enriched in the gene ontology (GO) annotations of “tissue development”, “cell adhesion”, and “cell differentiation” (Figure 3F), participated in the top kyoto encyclopaedia of genes and genomes (KEGG) pathway classes “infectious diseases”, “cancers”, “digestive system”, “neurodegenerative diseases” and “cellular community” (Table S2). While under the LPS stimulation for 12 h, 242 genes altered between shMEF2D and NC stable BV2 cells (Figure 3E), enriched in the GO annotations of “response to stimulus”, “immune system process” and “innate immune response” (Figure. 3F), involved in the top KEGG pathway classes of “immune system”, “infectious diseases” “cancers” and “signal transduction” (Table S2). These results strongly support the fact that MEF2D could impact the immune response and that knockdown MEF2D could change the immunoregulatory function of BV2 microglia cells.
Knockdown MEF2D significantly blocked the interferon response system
Interferon signal pathway is an essential part of the innate immune response in microglia by participating in the immune response to a variety of stimuli from endogenous and exogenous, triggering the phenotypical plasticity and the cascade of interferon-stimulated genes (ISGs). Interferons (IFNs) have been implicated in the dysregulation of immune responses in autoimmune diseases (20) and neurodegeneration (21, 22). Strikingly, we found that part of the down regulated genes of shMEF2D cells compared to NC, after LPS treatment for 12 h, were clustering to the interferon signal pathway, such as Cxcl10, Ifit1, Il12b, Il6, IRF7, Isg15, Mx2, etc. (Figure 4A). And most of the suppressed ISGs were induced mainly by IFN-Is pathway. These data well-documented that knockdown MEF2D obviously restrained the interferon response system. In other words, MEF2D is an indispensable element for the fully activation of interferon signaling pathway.
IFN-Is are transcriptionally regulated (23), and induced following recognition of pathogen components during infection by various host pattern recognition receptors. IRF7 and IRF3 are the most important transcriptional factors of the initiation of IFN-Is (24). The Genes network indicated that IRF7 is the key transcriptional factor of the significantly and differently expressed genes in shMEF2D and NC stable BV2 cells with LPS treatment (Figure 4B). The transcript abundance (in Read count) of IRF7 and IRF3 were evaluated, and the data showed that the mRNA level of IRF7 was suppressed, while IRF3 was not affected (Figure 4C). Also, the transcript abundances of IFN-Is such as IL-6, IL12B and CXCL10, were significantly suppressed by shMEF2D under LPS stimulation (Figure 4D). As IRF7 is a master regulator not only in the stage of initiation, but also in the second wave of cascade (24, 25), the data of RNA-seq make us to speculate that IRF7 may be the key factor for MEF2D to regulate the IFN-Is.
Transcriptional regulation of RSAD2 and IRF7 by MEF2D
To readout the key regulatory point of MEF2D, the down regulated genes in shMEF2D cells were screened with a putative MEF2 binding motif in the promoter sequences. Several genes including IRF7 were predicted to possess the potential sites (Figure 5A). To assess the direct binding of MEF2D, we carried out ChIP-PCR assay. The data showed that MEF2D bound specifically to the regions of the promoters of IRF7 that contains the putative sites by PCR. And the binding quantity was significantly induced in response to LPS treatment in BV2 cells (Figure 5B and C). We also performed qPCR to conform the subsequent IFN-Is. The data showed that LPS stimulation could significantly trigger IFN-I signaling pathway, and knockdown MEF2D markedly inhibited the elevation of IRF7, IFNA1, MX2, IFIT1, IFI27, ISG15, RSAD2, ZBP1 and DDX58 (Figure 5D). Together, these findings demonstrated that MEF2D directly regulate the transcriptional activity of IRF7, the key regulatory factor of IFN-Is (26, 27). These results strongly support the conclusion that MEF2D is a powerful regulator of the immune system, mainly through the IFN-I signaling pathway in BV2 microglia cells.
{kind=link}