Aplicability of molecular, cytometric and morphometric approaches in hybrid identification
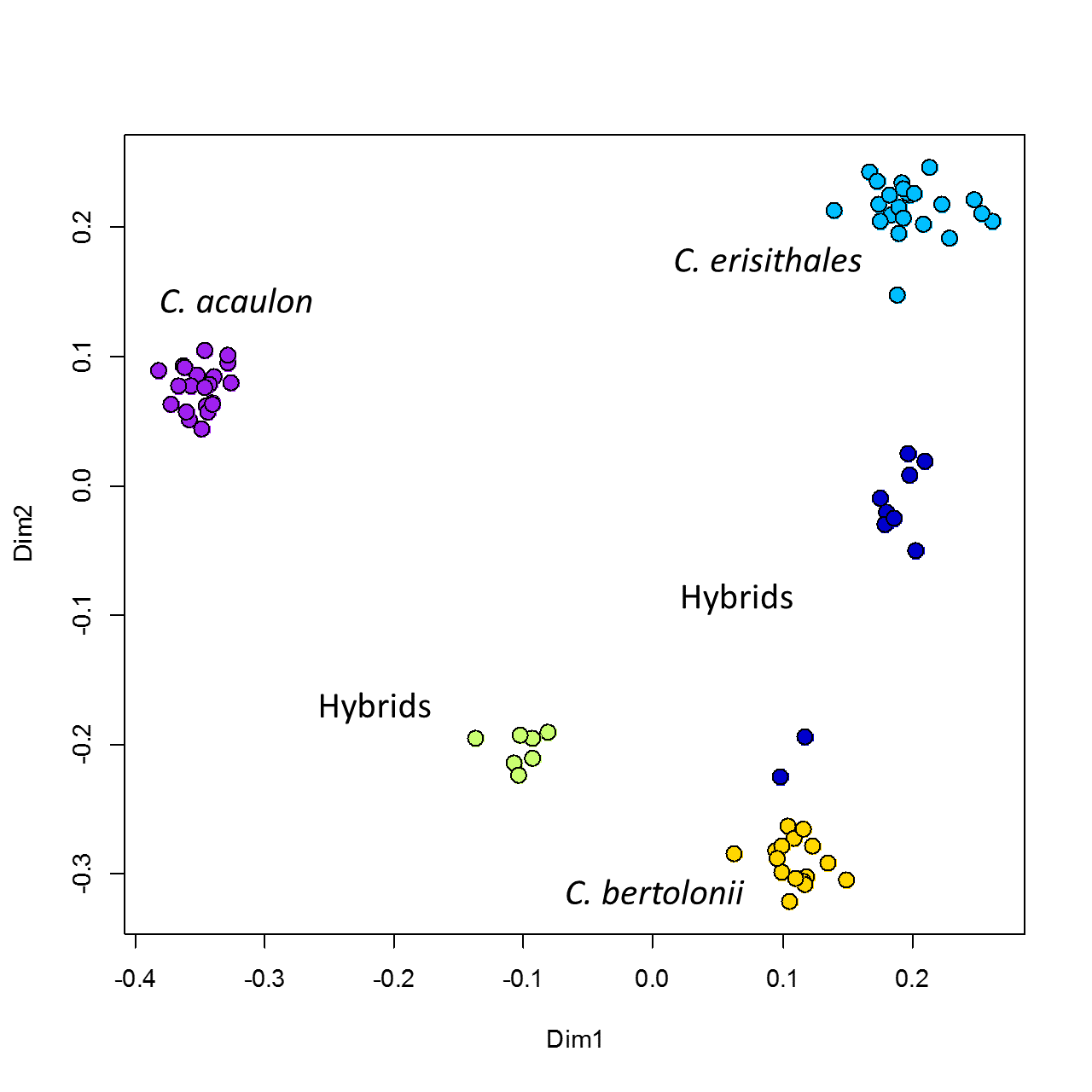
Although AFLP is currently considered to be rather outdated and is slowly taken over by NGS markers allowing the analyses of tens to hundreds of thousands of markers (Wang 2014), in our study it undoubtedly demonstrated the hybrid nature and parental affiliation of samples of Cirsium acaulon × C. bertolonii and Cirsium bertolonii × C. erisithales, as within the STRUCTURE analysis hybrids contained admixture from both parental species (Fig. 4) and distance-based analyses positioned them between the respective parents (Fig. 5, Suppl. Fig. S3). The STRUCTURE analysis worked well not only for the three parental species and the hybrids, but also when including 7 more closely related species occurring in Tuscany. The 10 well delimited STRUCTURE entities (K = 10, Suppl. Table S2) correspond to 10 species included in our study, which would indicate that despite frequent interspecific hybridization in this genus, none of these species is of hybrid origin.
Genome size estimation via flow cytometry indicated that the hybrids are diploid, which is the most common ploidy level in the genus (detected in nearly 80% of Cirsium species in Europe and Asia – Bureš et al. 2004, 2018). Flow cytometry was not a suitable tool for detecting the parental affiliation of either of the two hybrids (when statistical significance > 0.05 was taken into account). This is primarily given by the small differences between the hybrids and respective parents due to low variation on genome size across the genus (Bureš et al. 2004, 2018; Michálková et al. 2018) that is in an order of magnitude comparable to the error of individual cytometric estimation. Although the genome sizes and GC contents of hybrids were positioned more or less between the respective parental values, the difference was not always significant (Tukey HSD test; Fig. 6A, B), that is a consequence of the limited number of analyzed individuals and would likely be overcome by a larger number of measurements
Morphometric analysis detected some characters well delimiting respective parental species pairs and their hybrids (first five / seven characters in Tables 1A or B, respectively). Although the hybrids were usually positioned between the respective parents, they were usually shifted towards either of the parents (second and third sections in Tables 1A and B) or occasionally the hybrids were even out of parental range (e.g., C. acaulon × C. bertolonii has smaller number of secondary leaf lobes and more spiny leaves – Suppl Fig. S4N, X – and C. bertolonii × C. erisithales has narrower involucres, larger number of capitula and shorter distance from capitulum to 3rd stem leaf, than any of parents – Suppl Fig. S4C, H, I). The achenes of C. acaulon × C. bertolonii were also smaller than of any of the parents, nevertheless, they were not ripe and cannot be considered out of the parental range. This situation well illustrates that hybrid plants are more frequently a mosaicism of parental features rather than their average.
Gynodioecy of C. bertolonii and fertility of its hybrids
The hybrids of C. acaulon × C. bertolonii, similarly as two of nine individuals of C. bertolonii × C. erisithales (BERT×ERIS-ATP2c, BERT×ERIS-ATP4a) did not contain developed pollen in their anther tubes. One would easy interpret it as hybrid sterility. However, these individuals were actually females whose anther sterility has been inherited from the parental species – C. acaulon, C. bertolonii and C. erisithales – which all they are gynodioecious, i.e., containing hermaphrodites and females (= individuals with hermaphrodite flowers but sterile rudimentary anther tubes (synantherium – Suppl. Fig. S1) regularly in their populations. Similar co-occurrence of females and hermaphrodites (with viable pollen) was previously detected in hybrids of C. acaulon (with C. canum, C. oleraceum or C. pannonicum) and in hybrids of C. erisithales (with C. heterophyllym, C. oleraceum or C. palustre) by Bureš et al. (2010). The gynodioecious sexual dimorphism recognized in 275 (~ 2%) of angiosperm genera (Dufay et al. 2014) has so far been found in more than 20 species European species of Cirsium sect. Cirsium (Delannay 1978; Bureš et al. 2010). In our study, the gynodioecy of C. bertolonii is reported for the first time.
Besides the sterility/fertility of anther tubes, the conspecific females and hermaphrodites in Cirsium can also differ in other generative features (Bureš et al. 2018) as also demonstrated in our study for all three analyzed species (C. acaulon, C. bertolonii and C. erisithales) whose hermaphrodites have longer corollas, pappus but smaller achenes than females (Fig. 8; Suppl. Fig. S4A). Although larger flowers of hermaphrodites in most gynodioecious species are well known to evolutionary biologists from the time of Charles Darwin (1877), this fact is often ignored in taxonomic studies and determination keys. In Cirsium, especially among closely related species, the differences between the females and hermaphrodites within the species could even be higher that between the species as demonstrated for closely related C. greimleri and C. waldsteinii by Bureš et al. (2018). This is, fortunately, not the case in our recent study where the differences among C. acaulon, C. bertolonii and C. erisithales are rather larger than between genders within species (Fig. 8; Suppl. Fig. S4A), but for hybrid delimitation against the parental species the comparison of females with females or hermaphrodites with hermaphrodites is more reliable than when not considering the gender (Fig. 8).
The analyzed hybrids of C. bertolonii with C. acaulon at Monte Sagro did not contain ripe achenes which one would interpret as a hybrid sterility. However, in Cirsium, the absence of ripe fruits or low fruit fertility is relatively common even in regular non-hybrid plants (Bureš, pers. observ.). Therefore we do not consider undeveloped achenes in C. acaulon × C. bertolonii to be the evidence of reproductive barrier between these species via hybrid sterility. Fertility of C. acaulon hybrids with other Cirsium species has previously been confirmed e.g., by Pigott (1968), or by Bureš et al. (2010). In C. bertolonii × C. erisithales hybrids from Alpe Tre Potenze well developed achenes were detected suggesting the hybrids might be fertile as was demonstrated for some other hybrids of C. erisithales (Bureš et al. 2010).
The hybrid samples from ATP1 locality in the Alpe Tre Potenze were situated outside the cluster of hybrids C. bertolonii × C. erisithales towards the first of parents in STRUCTURE (Fig. 4) or in distance-based analyses (Fig. 5; Suppl. Fig. S3), similar shift of these samples toward C. bertolonii can be seen in genome size and genomic GC content (Fig. 6A, B); in morphometric analysis such shift is more or less present in majority of characters except three characters related to leaf lobes where the samples from ATP1 were strongly shifted towards C. erisithales or even exceeded it that finally resulted into the absence of the shift to C. bertolonii in CDA (Fig. 9) otherwise observable in other analyses. One could speculate that this pattern resulted from backcrossing (of C. bertolonii × C. erisithales with C. bertolonii), however, it can also be the case of F2 resulting from random segregation of chromosomal homologs during gametogenesis, and selfing of homoploid F1 hybrid (or outcrossing between F1 hybrids prevailing in analyzed populations in Alpe Tre Potenze). In any case, the occurrence of such genetically shifted individuals supports the fact that primary hybrids (F1) of C. bertolonii × C. erisithales may be at least partially fertile as documented for other Cirsium hybrids (e.g., by Correns 1916; Bureš et al. 2010; Michálková et al. 2018).
The possible threat of genetic erosion of C. bertolonii
Somatic DNA content estimated for C. bertolonii in our study is very similar to DNA contents of other diploid Cirsium species (Bureš et al. 2004, 2018; Michálková et al. 2018) and chromosome number 2n = 34 based on achenes sampled from the individuals in the type locality Monte Sagro also undoubtedly confirmed the diploid status of C. bertolonii as previously suggested Miceli and Garbari (1976). Therefore, previously reported 2n = 12 (Garbari 1970) needs to be considered erroneous and neither different ploidy level nor some kind of chromosomal rearrangement do not protect C. bertolonii against hybridization with other frequently hybridizing and predominantly diploid species of Cirsium sect. Cirsium as it is the case for rare mountain endemic tetraploid (2n = 68) C. appendiculatum in Balkan Mountains or C. waldsteinii in the Carpathians (Bureš et al. 2018) Our study also confirmed that C. bertolonii really can produce (at least partially) fertile hybrids with two diploid relatively more abundant and more widespread congeners: C. acaulon and C. erisithales – C. acaulon (sensu stricto) occurs almost continually in Western and Middle Europe – from England, France and Northern Spain to Southern Sweden, the Baltics, Poland, Czech Republic, Austria, Italy, Slovenia, Croatia, Bosnia and Herzegovina, with dispersed occurrences also in Slovakia, Hungary and Romania and C. erisithales is growing in most of the higher European mountains (or their parts) such as in Easternmost Pyrenees, Massif Central, the Alps, Northern and Central Apennines, the Dinarides and the Carpathians (Meusel and Jäger 1992). Across their nature ranges, both species willingly hybridize with most of their sympatric congeners (Pignatti 2018; Talavera 2017; Stace et al. 2015; Bureš 2004; Wagenitz 1987; Nyárády 1964) therefore, it is not surprising that they hybridize when co-occurring with the rare C. bertolonii with whom they share ecological preferences to calcareous substrates (Ferrarini 1967; Gabellini et al. 2006; Tomaselli et al. 2019).
In the area of C. bertolonii distribution, C. acaulon is scattered in both the Apuan Alps and Tuscan-Emilian Apennines (Bertoloni 1819; Caruel 1860; Archbald 1874; Gibelli and Pirotta 1882; Ferrarini 2000; Romagnoli and Foggi 2005; Venturi 2006; Pierini and Peruzzi 2014) while C. erisithales seems to be common only in the Tuscan-Emilian Apennines (Caruel, 1860; Archbald 1874; Gibelli and Pirotta 1882; Venturi 2006; Foggi et al. 2007; Pierini and Peruzzi 2014) and in the Apuan Alps it is probably restricted only to two localities – Monte Contrario (south of Minucciano) and Col di Favilla (north of Stazzema) (Ferrarini 2000; Pierini and Peruzzi 2014). The uneven distribution of C. acaulon and C. erisithales is rather reflected by hybridization observed in population of C. bertolonii. In Monte Sagro (Apuan Alps) where abundant populations of C. acaulon and C. bertolonii are intermingled, their hybrids occur scarcely. In Alpe Tre Potenze (Tuscan-Emilian Apennines), we did not find a “pure” C. bertolonii as all sampled individuals were hybrids or genetically eroded (Fig. 4, 5). Since C. spinosissimum is absent in the Apuan Alps (Zanotti and Cristofolini 1985; Ferrarini 2000), the previous report on the hybrid C. erisithales × C. spinosissimum by Caruel (1865) – from the vicinity of Minucciano – needs to be ascribed to C. bertolonii [= C. spinosissimum var. bertolonii sensu Caruel, l.c.] × C. erisithales as also concluded by Pierini and Peruzzi (2014). The hybrid has therefore found by Caruel (1865) near of one of the two localities where C. erisithales occurs in the Apuan Alps – in northern slopes of Monte Contrario Mt. south of Minucciano (Ferrarini 2000). It should be noted that also relatively close to the second locality of C. erisithales in the Apuan Alps – Col di Favilla (Ferrarini 2000) – there was also in 19th century collected a herbarium specimen that is morphologically very similar to the plants identified as a hybrid of C. bertolonii × C. erisithales in our study (“Sopra Campanice nelle Alpi Apuane” [= above the settlement of Campanice], sine collectore 27th July 1863, MNHN-P-P04316612), further probably hybrid plant C. bertolonii × C. erisithales has been collected in close locality south of this second occurrence at “Südeingang des ersten Strassentunnels, bei der Abzweigung nach Arni, sm. 1050 m by C. Simon (26th. June 1957 BASBG-00025459, BASBG-00025460).
The higher prevalence of hybrid between C. bertolonii and C. erisithales may also be due to overlap in their flowering times during Late Spring–Early Summer; whereas C. acaulon blossoms later – in Summer–Early Autumn, depending on altitude (Michálková and Bureš pers. observ.). The phenological difference of C. acaulon was notable during our repeated visits to Monte Sagro – in July, C. bertolonii was in pollination optimum, whereas C. acaulon had only developed inflorescence buds and the hybrid already had capitula, but was not flowering yet; in October the shoots of C. bertolonii were almost faded, whereas some of the hybrids were still blossoming as well as C. acaulon.
Scattered distribution of hybrids C. acaulon × C. bertolonii in Monte Sagro and the exclusive presence of C. bertolonii × C. erisithales in four studied populations in Alpe Tre Potenze suggest that the hybridization of C. bertolonii with both these species is repeatedly ongoing which is further confirmed by abovementioned herbarium specimens and literary report as well as by an older herbarium specimen of C. acaulon × C. bertolonii collected probably by A. Bertoloni in Catino dell Sagro in Northern slope of Monte Sagro in the beginning of 19th century and later deposited in the Viennese Natural History Museum (Bertoloni s. a. W 1929/1457; Suppl. Fig. S5). From a taxonomic point of view, it is somewhat worrying that hybridization with C. acaulon takes place directly on the locus classicus of C. bertolonii – within the Monte Sagro population. However, based on Bertoloni´s “Amoenitates“, we can conclude that both species coexist here for at least 200 years without any of them getting extinct (Bertoloni 1819: 404–405); the neotype from this locality was selected by Zanotti and Cristofolini (1985: 35; http://parlatore.msn.unifi.it/img72/FI001979.jpg) does not evince any sign of hybridization.
Although, the situation in Monte Sagro illustrates a relatively long coexistence of C. bertolonii with C. acaulon, repeatedly ongoing hybridization, fertility of hybrids C. bertolonii with C. erisithales, as well as overlap in blossoming and ecological preferences could pose a serious threat for genetic integrity of the rare mountain North Apennine endemic C. bertolonii particularly by genetic erosion from C. erisithales in Alpe Tre Potenze or in the whole Tuscan-Emilian Apennines – similarly as for other rare diploid European narrowly distributed high mountain thistles, such as C. alsophilum, C. greimleri, C. carniolicum or C. rufescens (Bureš et al. 2018; Michálková et al. 2018).
Taxonomic Conclusion – Description Of The Hybrids
Morphological intermediacy of the hybrids and their differences against parental species are best fitted by the features which variation is described in Tables 2A, B. Based on the morphological intermediacy of the found hybrid individuals (Figs. 2, 3, Suppl. Fig. S2) supported by molecular and cytometric analyses (Figs. 5 and 6), their repeated origin and the presence in herbarium collections, there are two new nothotaxa that can be recognized:
Cirsium ×sagrense Michálková et Bureš, nothosp. nova
= C. acaulon (L.) Scop., Annus Hist.-Nat. 2: 62, 1769 × C. bertolonii Spreng., Syst. Veg., ed. 16. [Sprengel], 3: 377, 1826
Diagnosis: The hybrid differs from both parental species in its pinkish white corollas and is intermediate between them in stem length – 22–28 cm [1–5 (–35) cm in C. acaulon, 45–60(–75) cm in C. bertolonii], in pappus length 19.8–21.2 mm [(21.4–) 22.1–27.5 (–29.5) in C. acaulon, (14.6–) 17.9–20.9 (–21.7) in C. bertolonii], in (1–) 2–5 capitula – [1–2 (–5) in C. acaulon, 2–8 in C. bertolonii], in 2.5–3 (–3.4) cm distance between lateral lobe apices in basal leaves [(0.8–) 1–2.2 (–3.0) cm, in C. acaulon, 3.2–4.4 cm in C. bertolonii]. From C. acaulon the hybrid differs in the hairy abaxial surface of involucral bracts (glabrous in C. acaulon), in longer (> 0.5 mm) terminal spines of outer and middle involucre bracts [either absent or very short (< 0.5 mm) in widespread C. acaulon subsp. acaulon but could be longer than 0.5 mm in Iberian endemics C. acaulon subsp. gregarium (DC.) Talavera], and in presence of uppermost subtending bracts (absent in C. acaulon). From C. bertolonii the hybrid differs in the densely wooly indumentum with undulated longer (> 1 mm) hairs [C. bertolonii has sparse indumentum with hairs shorter than 1 mm], in middle leaves that are 3.3–6.7-times longer than wide (C. bertolonii have middle leaves 1.1–3.2-times longer than wide), in involucres 1.5–1.8 times longer than wide (C. bertolonii have involucres 0.8–1.6 longer than wide); the indumentum of abaxial surface of involucral bracts is sparser and formed by non-glandulous hairs in the hybrid (denser, with glandulous hairs in C. bertolonii); terminal spines on outer and middle involucre bracts of the hybrid are shorter (0.5–1 mm) than those of C. bertolonii (> 1 mm).
Type: Italy: Apuan Alps; Vergheto: grassland slope between A Foce Pianza and Monte Sagro, 2 km SSE of the village; 44°06'40.3"N, 10°09'20.6"E; altitude 1496 m a.s.l.; coll. E. Michálková & S. Němejc, 6/Oct/2018 (holotype: BRNU; isotypes: PR; WU).
Description: Perennial spiny plant, 22–28 cm tall. Stem erect, rather branching on the top, unwinged, with dense wooly indumentum particularly in lower part formed by undulated hairs longer than 1 mm. Leaves oblanceolate, pinnately cleft–parted, spinous at the margins, densely wooly at abaxial side; bottommost leaves 21–30.2 cm long and 3.8–6.3 cm wide, middle 9.5–15 cm long and 1.8–3.7 cm wide; stem leaves gradually transiting to subtending bracts; uppermost subtending bracts narrow (2–10 mm wide), with perpendicularly protruding spines. Capitula (1–) 2–5 on separate peduncles. Involucre ovoid, 12.1–28.1 mm wide (incl. protruding ends of involucral bracts); involucral bracts ovate-lanceolate, outermost short, glabrous on the outer (abaxial) surface, rarely with hairs on midrib in upper part, outer and middle with short (0.5–1 mm) terminal spine, middle 1.8–2.8 mm wide. Corolla pinkish white, 23.1–24.3 mm long. Pappus 19.8–21.2 mm long. Achenes dark brown, (3.8–) 4.3–4.9 mm long (not well developed). Flowering from late July until early October.
Etymology: The epithet is derived from the mountain Monte Sagro, where the hybrids between C. acaulon and C. bertolonii were found.
Cirsium × abetonense Michálková et Bureš, nothosp. nova
= C. bertolonii Spreng., Syst. Veg., ed. 16. [Sprengel], 3: 377, 1826 × C. erisithales Scop., Annus Hist.-Nat. 2: 62, 1769
Diagnosis: The hybrid is intermediate between its parents by corolla length 17.1–20.4 mm [(12.8–) 13.6–17.6 (–17.9) mm in C. erisithales, (20.8–) 21.5–24 (–24.6) mm in C. bertolonii], by pappus length 15.6–18.7 mm [(11.7–) 12.0–16.2 (–17.1) mm in C. erisithales, (14.6–) 17.9–20.9 (–21.7) mm in C. bertolonii], in 2–7.4 (–9) mm long terminal spines of middle lobes of middle leaves [0.2–1.5 (–1.8) mm in C. erisithales, 3.3–10.3(–14) mm in C. bertolonii], and in sparse glandular abaxial indumentum of outermost involucral bracts – [dense glandular in C. bertolonii, glabrous in C. erisithales]. From C. erisithales the hybrid differs by achene length 5–5.7 mm [(3.6–) 4–4.9 (–5.1) mm in C. erisithales], in presence of leaves in the upper part of the stem, in presence of narrow uppermost subtending bracts, and presence of 0.5–1 mm terminal spine of outer and middle involucral bracts [in C. erisithales is absent or very short (< 0.5 mm)]. From C. bertolonii the hybrid differs in deeply lobed basal leaves (entire or shallowly lobed that are in C. bertolonii), shorter (< 1 mm) terminal spines of outer and middle involucral bracts (> 1 mm in C. bertolonii), and by (1–4) narrow uppermost subtending bracts (> 5 in C. bertolonii).
Type: Italy: Tuscan-Emilian Apennines; Abetone: N slopes of the mountain range, 800 m S of Val di Luce; 44°7'20.4"N, 10°37'40.2"E; altitude 1677 m a.s.l; coll. P. Bureš & E. Lajkepová, 5/Jul/2017 (holotype: BRNU; isotypes: PR; WU).
Description: Perennial spiny plant, 40–95 cm tall. Stem erect, branching, sometimes nodding on the top, unwinged, with pubescent indumentum particularly in lower part and below the capitula. Leaves oblongate, pinnately divided or even pinnatisect, spinous at the margins with sparse hairs on abaxial surface gradually transiting to subtending bracts; bottom leaves (40.2–) 40.4–57.3 (–60.3) cm long, (15.1–) 15.8–20.6 (–21.6) cm wide; middle leaves (16.6–) 20.1–38.3 (–41.2) cm long, (9.3–) 9.6–17.2 (–20.7) cm wide. Uppermost subtending bracts narrow (1–7 mm wide) with sparse glandular hairs on the margin and perpendicularly protruding shorter spines. Capitula 3–7 (–9) in clusters or on separate peduncles. Involucre ovoid, 20.1–40.9 mm wide (incl. protruding ends of involucral bracts); involucral bracts ovate-lanceolate, outermost short with sparse glandular hairs on the outer surface, outer and middle involucral bracts with short (0.5–1 mm) terminal spine, middle 1.5–2.2 mm wide. Corolla yellowish, 17.1–20.4 mm long. Pappus 15.6–18.7 mm long. Achenes light brown, 5–5.7 mm long. Flowering in July and August.
Etymology: The epithet is derived from the town Abetone in proximity of Alpe Tre Potenze, where the hybrids between C. bertolonii and C. erisithales were found.
{kind=link}
{kind=link}
{kind=link}