Male and female MSCs have similar growth and phenotypical characteristics
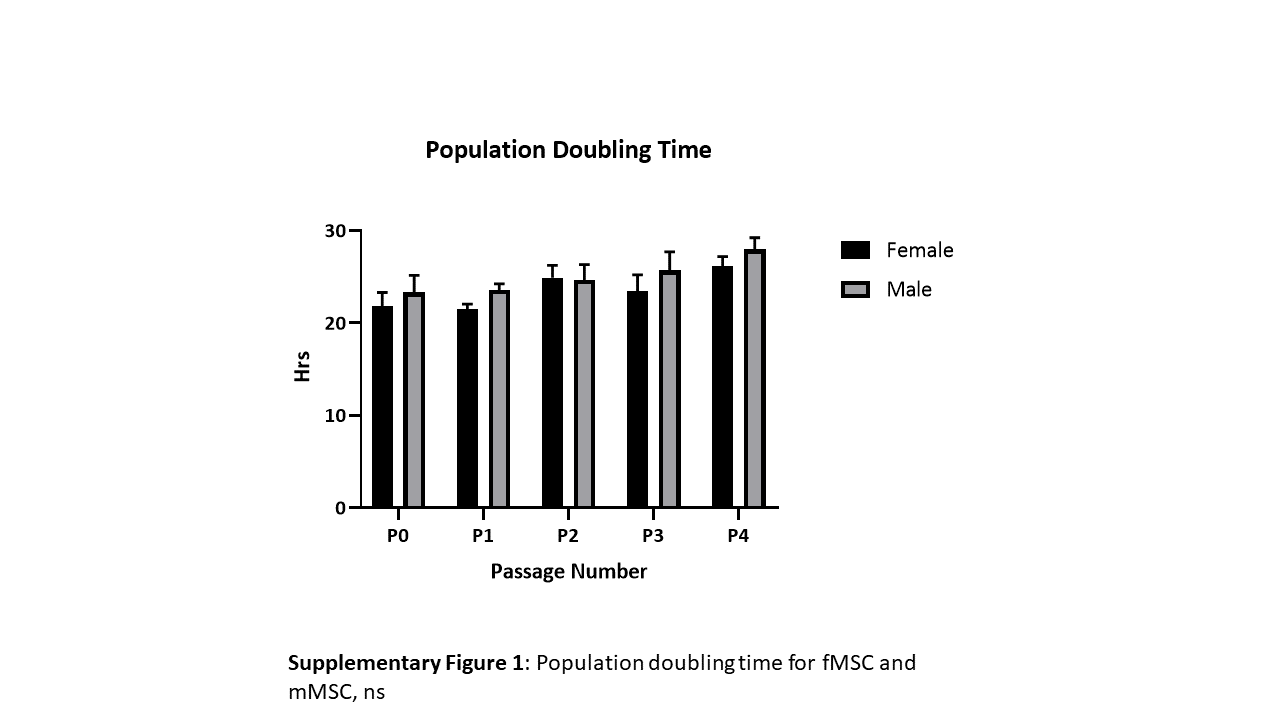
MSC doubling per day and total doublings showed no significant differences between the donors up to passage 4 (supplementary Fig. 1) and all MSCs showed a fibroblastic like morphology (Fig. 1).
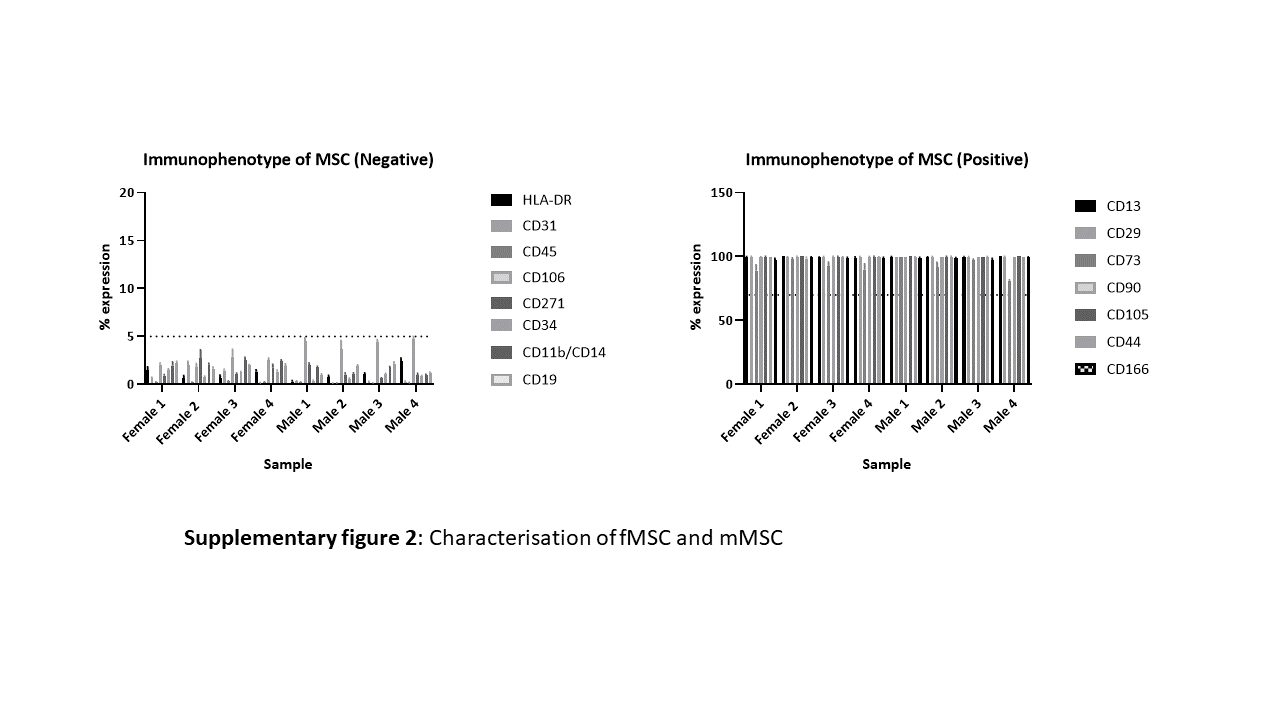
Additionally, male and female showed cell surface expression patterns consistent with MSC characterisation [9][11] Positive (≥ 70%) for classical MSC markers CD90, CD105, CD73 and negative for CD45, CD34, CD11b, HLA-DR and CD19. In addition, in an extended MSC characterisation panel all MSC were positive for CD166, CD44, CD13 and CD29 and negative (≤ 5%) for CD31 and CD271 via FACS (Supplementary Fig. 2).
Female MSCs have greater immunosuppressive properties than male MSCs
To test for difference in the immunosuppressive properties of MSCs derived from males (mMSC) and females (fMSC), MSCs were used to alter PBMC proliferation. Isolated male and female PBMC were stained with CellTrace CFSE and subsequently stimulated with MACSiBead anti-CD2, anti-CD3, anti-CD28 microparticles. PBMC from both male and females were cultured with primed male and female MSCs and were analysed on FACSCalibur for immune suppression. Figure 2 shows representative histograms showing the effect of fMSC (Fig. 2a) and mMSC (Fig. 2b) on PBMCs. The greater the loss of fluorescence, the greater the amount of proliferation. The suppression of PBMCs proliferation is represented by a shift in fluorescent intensity toward the unstimulated control. All fMSCs showed a more pronounced shift toward unstimulated controls when combined with PBMCs relative to mMSCs (Fig. 2a and b). Moreover, fMSC donors consistently presented a higher suppression rate than mMSC (Fig. 2c). The suppression rates were significantly higher (p < 0.0001) when fMSC were used to suppress both male and female PBMC (60.7+/-15.6 and 67.9+/-10.4), relative to mMSC (22.5+/-13.6 and 29.4+/-9.3) (Fig. 3). This data suggests increased MSC mediated suppression of PBMCs by fMSC compared to mMSC is entirely due to the competency of fMSC.
Male and female MSCs respond similarly to inflammatory mediators.
In addition to T-cell inhibition via paracrine mediators, it has long been recognised that cell surface markers including iCAM-1, VCAM-1 and PD-L1 play essential roles in MSC potency through their role in mediating cell-to cell contact [39–43]. Since primed fMSC have been shown to have a greater immune suppression than primed mMSC, this prompted us to ask whether the MSC response to inflammation was sex specific. To test this, we assessed the expression of iCAM-1, VCAM-1 and PD-L1 together with Interferon Gamma Receptor 1 (IFGR1) HLA-DR, HLA-ABC, HLA-G and HLA-E.
When MSC were not exposed (unprimed control) to inflammatory cytokines, there was no difference in the level of expression of iCAM, NT5E, HLA-ABC, PD-L1, HLA -DR, HLA-G, HLA-E or VCAM-1 (Fig. 4a).
When MSC were primed by exposing them to inflammatory cytokines, IFN-γ and TNF-α, (Fig. 4b) all donors upregulated VCAM-1 (fMSC 85.5+/- 16, mMSC 80.27 +/- 15.4), iCAM-1 (fMSC 99.77 +/- 0.2, mMSC 99.14 +/- 2.1), PD-L1 (fMSC 99.3+/-0.85, mMSC 98.8+/-2.05), HLA-DR (fMSC 64.1 +/- 26.2, mMSC 70.1 +/- 32), HLA-E (fMSC 78.8 +/- 10.4, mMSC 73.04 +/-25.7) and NT5E (fMSC 95.9% +/- 3.8. mMSC 93.1 +/- 3.6). In contrast, there was no effect on the expression level of HLA-G and IFNGR1. HLA-G remains virtually undetected (< 5%) across all samples tested.
This indicates that all MSC donors irrespective of sex, possess similar ability to regulate cell surface markers when presented with an inflammatory microenvironment.
Post Primed female MSCs sustain VCAM-1 while male MSCs sustain HLA-DR
Since there was no difference in the induction of cell surface receptor expression between male and female MSC, this prompted us to investigate how mMSC and fMSC respond post inflammation. MSCs were primed for 24 hrs and cultured for a further 24hrs in MSC culture media. MSCs were assessed for immunoregulatory cell surface markers. Figure 4(c-e) shows fMSC sustain the presence of VCAM-1 (p = 0.002) more so than mMSC, while mMSC sustain HLA-DR expression more than fMSC (p = 0.04). There was no significant difference in the expression of PD-L1, iCAM, IFNGR1, HLA-ABC, HLA-G, HLA-E and NT5E 24 hrs post priming.
Male and female MSCs selectively regulate CD8, CD25 and CD69 expression in activated PBMCs.
It is well known that MSC interaction with CD4+T cells is imperative to immunomodulation. MSCs alter T-cell phenotypes from an inflammatory to anti-inflammatory phenotype [44, 45]. To investigate whether inherent differences exist between male and female PBMCs and/or how MSCs interact with PBMCs and alter their phenotype, we assessed cell surface markers of PBMCs alone (Resting and Activated) and after 6 days in co-culture with primed MSCs.
Analysis of the lymphocyte population showed no differences in the level of expression of CD3, CD4, CD8, IL-2 receptor CD25 or the early activation marker CD69 between male and female PBMCs when resting, activated with MACSiBead microparticles for 3 days (early activation) or after 6 days stimulation (supplementary Fig. 3). CD8 and CD25 were upregulated in response to MACSiBead microparticle stimulation. CD8 and CD25 were increased after 3 days and remained high at day 6 (Fig. 5). In contrast, CD69 expression showed an early activation peak which declined by day 6 (resting 0.8%, 3 days activated 38.1% and 6 days activated 6.7%, Fig. 5).There was no change to CD19+ B cell numbers in response to MACSiBead microparticles (data not shown)
When activated PBMC were co-cultured with either mMSC and fMSC for 6 days (Fig. 5),, the expression of CD8 (Fig. 5a, 6d) and CD25 (Fig. 5b, 6e) was downregulated relative to activated PBMC without MSC. The sex of the MSCs dictated the level of suppression, mMSC were less effective at downregulating CD8 in female and CD25 in male and female PBMCs than fMSC (p < 0.05). More over, in the presence of fMSCs, but not mMSCs, CD69 expression in PBMC from both sexes was sustained for 6 days (Fig. 5c and f), indicating an MSC sex related difference in mediating PBMC cell surface marker regulation.
Primed female MSCs produce higher levels of IDO, PGE-2 and IL-1RA but lower levels of G-CSF than male MSCs.
To further examine the immunomodulatory capabilities of male and female MSC, we compared the secretion characteristics of both sexes post culture.
1. Positive analytes
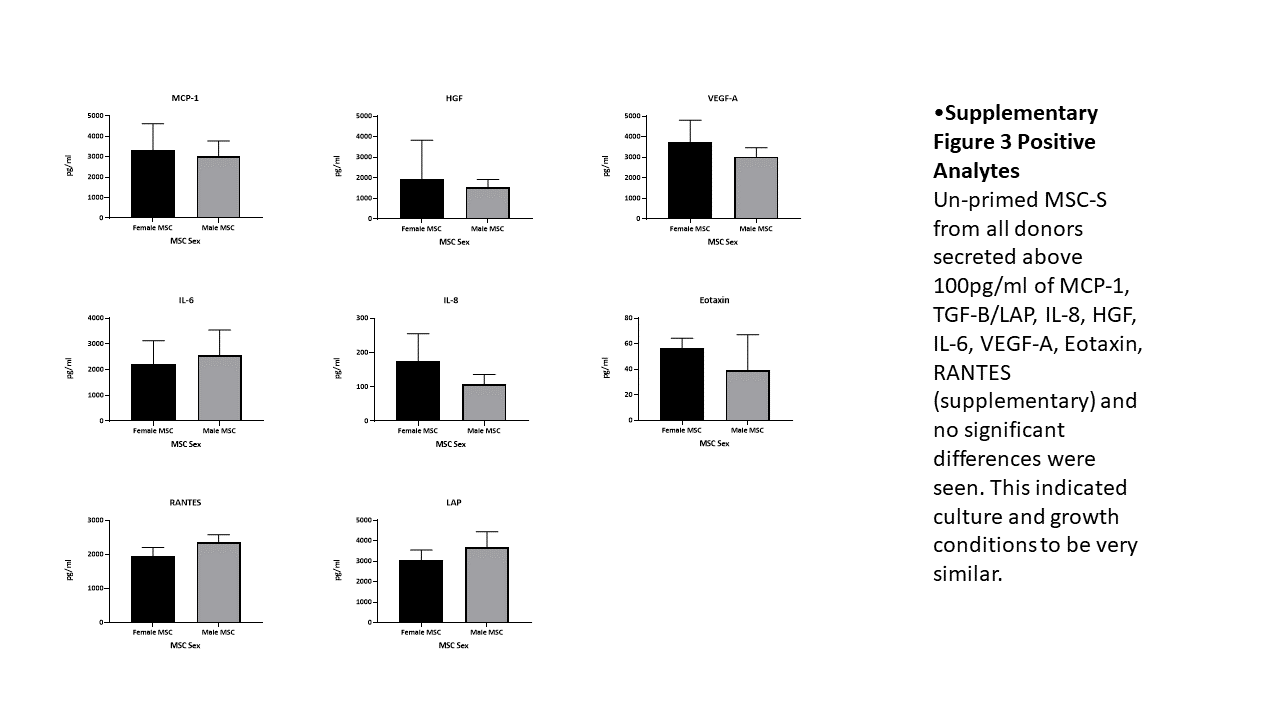
There were no significant differences for Unprimed MSC-S from all donors for MCP-1, TGF-B/LAP, IL-8, IL-6, VEGF-A, Eotaxin, RANTES (supplementary Fig. 3) indicating culture and growth conditions were similar irrespective of MSC sex.
2. Induced analytes
To gain further insight into the paracrine mechanisms of MSC immunomodulation, we also harvested the MSC-S 24hrs post priming and assessed protein expression. MSC-S from all donor’s post priming showed increased expression of MCP-1, TGF-B/LAP, IL-8, IL-6, VEGF-A, HGF, IP-10, MIP-1α and MIP-1β (data not shown). There was no significant difference in the level of HGF, MCP-1, VEGF-A, IL-6, TNF-α or IFN-y in the secretome from male and female MSCs (Fig. 6). In contrast, MSCs from all female donors secreted higher concentrations of IDO1 than their male counterparts (Fig. 7a 3301pg/ml vs 1699pg/ml respectively). Female activated MSC also showed increased expression of PGE-2 (Fig. 7c fMSC 6142pg/ml vs mMSC 2448 pg/ml) and the “IL-1 inhibitor”, IL-1RA (Fig. 7b fMSC 1025 pg/ml vs mMSC 701 pg/ml) whereas the opposite was seen for G-CSF (Fig. 7d fMSC 503 pg/ml vs mMSC 806 pg/ml). Analytes that were assessed but not detectable are indicated in the Supplementary section.
Inhibition of IDO function completely ablates the suppressive function of female MSCs.
IDO1 is responsible for the catabolism of Tryptophan, which is an essential part of T-cell activation and proliferation. Our data shows fMSCs secrete greater quantities of IDO1 than mMSCs. To determine whether IDO contributed to the efficacy of fMSCs, IDO was inhibited from fMSCs using Epacadostat/INCB24360 (EPA) a known inhibitor of IDO1 [46]. CFSE stained female PBMC were co-cultured with fMSC with and without EPA treatment and the level of suppression determined.
Unstimulated PBMCs had a CFSE binding mean fluorescence intensity (MFI) of 2059+/-249.5. Post stimulation, the PBMC MFI dropped to 70.52+/-8.2, representing cell division. When Primed MSC were added to the culture MFI readings were closer to an unstimulated state (1421+/-248.8) indicative of immune suppression, or impaired daughter cell division. However, when MSC were co-cultured with activated PBMC in the presence of EPA, the level of immune suppression was reduced from ~ 75–17% (Fig. 8a and b). Moreover, IDO1 expression by MSCs was not inhibited by EPA (Fig. 8c) rather EPA blocked IDO1 binding and inhibited the enzymatic activity necessary for T-cell activation.
To determine whether the expression of IDO1 correlated with PBMC suppression we carried out linear regression analysis of IDO levels from MSCs from males (n = 4) and females (n = 4) with their immunosuppression ability on activated PBMC. MSC immunosuppressive capacity significantly correlated with IDO1 expression levels (R2 = 0.77, Fig. 9). Potentially providing a method for determining MSC efficacy.

{kind=link}
{kind=link}
{kind=link}