Design and construction of the copper-inducible T7 RNAP system
The trace element of copper has important functions in bacteria [19–21], and its homeostasis in E. coli has been extensively studied [22, 23]. The copper homeostasis is a tightly regulated process, to maintain the physiological level of copper. There are three pathways contributing to the copper homeostasis in E. coli: (1) the Cue system regulates the cytosolic copper level, and the major components of CopA and CueO are regulated by CueR [24, 25]; (2) the Cus system regulates the periplasmic copper level, and the major components of the proton cation reverse transporter CusCBA and the periplasmic copper-binding protein CusF that are regulated by CusSR [26, 27]; and (3) the Pco system enables the E. coli with additional copper resistance [28, 29].
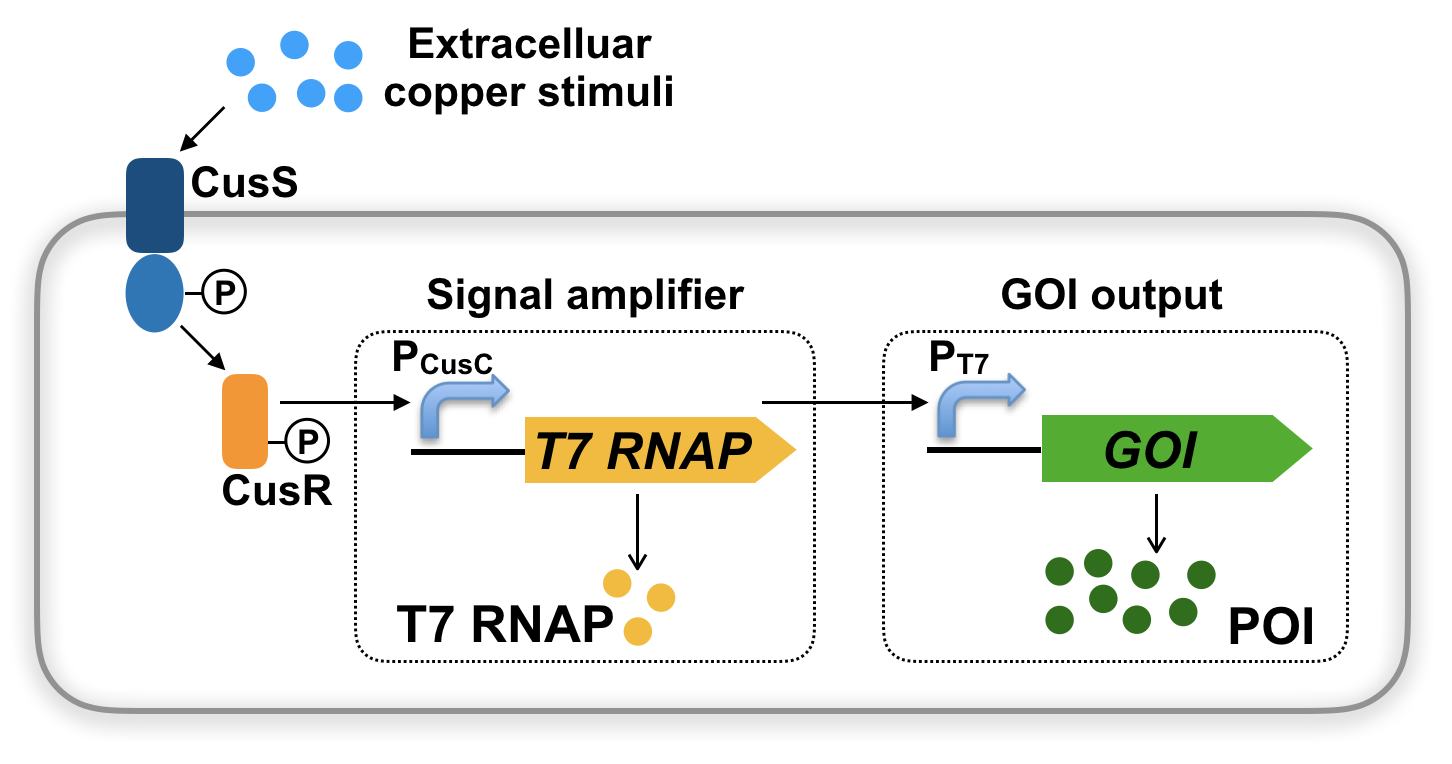
In this study, we chose the CusC promoter (PCusC) with high sensitivity [16, 17] to drive the T7 RNAP expression (Fig. 1). Further incorporating GOI under the PT7 promoter would make GOI expression to be triggered by copper induction (Fig. 1). In particular, the two-component system of CusSR responds to the extracellular copper stimuli and activates the PCusC, which makes the expression of T7 RNAP under the control of the PCusC, and finally T7 RNAP initiates the PT7 promoter to drive the expression of GOI. To construct the PCusC-T7 RNAP system, we used CRISPR/Cas9 technique to directly integrate T7 RNAP at the CusC locus of BW25113 (Fig. 2A), and the resulting strain was designated as BW-Cu.
To test the feasibility of copper-inducible T7 RNAP system, a lacO-omitted plasmid with eGFP reporter gene (pCDF*-eGFP) [13] was transformed to BW-Cu, and the resulting strain BW-Cu with pCDF*-eGFP was designated as BW-Cu-eGFP. When the strain BW-Cu-eGFP was treated with different concentrations (0, 1, 2.5, 5, 10, 15, and 20 µM) of Cu2+, we obtained dose-dependent expressions of eGFP in both glucose- and glycerol-containing medium (Fig. 2B). However, we found that the copper-inducible system functioned better in glycerol medium, and the maximum signal output could reach 5.6-fold higher than that in glucose medium. The induction ratio in glycerol medium was about 35-fold over that of the control without copper supplementation (Fig. 2C).
To further confirm whether the copper-stimulus was recorded into T7 RNAP expression for a prolonged gene expression, we decided to further investigate time course studies of eGFP expression. As shown in Fig. 2D, the eGFP signals were well maintained under the tested conditions, indicating that the signal of copper stimuli was well recorded to the secondary layer of T7 RNAP. Therefore, the layered genetic design did outperform the traditional PCusC-controlled GOI in regarding to the signal output and memory recording (Fig. 1).
Harnessing The Copper-inducible T7 Rnap System For The Pca Production
Next, we sought to explore the copper-induced gradient system for PCA production. As shown in Fig. 3A, PCA synthesis can be realized by connecting the endogenous shikimate pathway with the heterologous introduction of asbF encoding 3-dehydroshikimate (DHS) dehydratase (DSD) [30]. In this study, we introduced asbF from B. cereus together with the feedback resistant aroGfbr encoding 3-deoxy-d-arabinoheptulosonate 7-phosphate (DAHP) synthase [31] into the strain BW-Cu, and the resulting strain was designated as BW-Cu-PCA. When the strain BW-Cu-PCA was treated with three concentration gradients of Cu2+ (10, 15, and 20 µM), we found that all the strains with copper induction produced substantially higher amounts of PCA than that of the control without copper supplementation (Fig. 3B). The PCA was rapidly synthesized in the early 24 h, while further cultivation did not improve the PCA yields. In addition, 10 µM Cu2+ was sufficient to induce the cells to produce 16.81 mM PCA (2.59 g/L) and the PCA productivity was relatively stable maintained during the fermentation process. For unknown reasons, we found that the growth of PCA-producing strains was slightly higher than that of the control (Fig. 3B).
Further Improvement Of Pca Production Using Crispri Method
To further improve the PCA yield, we further attempted to redistribute the central metabolism toward PCA synthesis using the CRISPRi method (Fig. 3A). The gene targets of pykA, pykF, ppc, aroE, and ydiB were chosen to restrict the consumption of phosphoenolpyruvate (PEP) and DHS as previously reported [13]. Therefore, strain BW-Cu-PCA was further introduced with pCDF*-dCpf1-M3 (Table 2), and the resulting strain was named as BW-Cu-PCAM3. To remove the difference in plasmid burden caused by the addition of the plasmid, strain BW-Cu-PCA was transformed with the empty vector of pCDF* (BW-Cu-PCASm) to serve as a control. As shown in Fig. 3C, strain BW-Cu-PCASm afforded to produce a total of 9.2 mM PCA (1.42 g/L) after 72 h. In comparison, strain BW-Cu-PCAM3 with CRISPRi produced 24.35 mM PCA (3.75 g/L) after 72 h, which was almost 2.6-fold improvement over that of BW-Cu-PCASm. It should be noted that the introduction of CRISPRi module delayed the cell growth (Fig. 3C), although the strain could eventually reach the similar biomass. Nevertheless, we managed to efficiently improve the PCA titer using the CRISPRi technique.
Since the genes (pykA, pykF, ppc, aroE and ydiB) targeted by CRISPRi are involved in the central metabolism, we reasoned that the timing of copper addition is important to balance the growth and PCA production. We therefore verified the effect of different timings of copper addition (3 h, 6 h, 9 h, and 12 h). As shown in Fig. 3D, the highest yield of 26.76 mM PCA (4.12 g/L) occurred when Cu2+ was added after 6 h inoculation. Meanwhile, all groups eventually reached the similar biomass (Fig. 3D). However, we found that supplementation of copper at 12 h favored the biomass accumulation instead of PCA production. Therefore, the copper-induced gradient expression system was capable of controlling metabolic pathways in a temporal and dose-dependent manner.
{kind=link}