Numerous evidences suggest that obesity induced by high-fat diet can adversely af-fect the function of brain [36, 37] and a growing number of studies link gut dysbiosis to neurological and psychiatric disorders [38–40]. Our results demonstrated that different types of high dietary fatty acids might induce obesity, pathological changes in hippo-campal CA1 area, and alterations of gut microbiota in mice. Transplantation of fecal mi-crobiota shaped by multiple high dietary fatty acid diets, possibly through damage to the hippocampus CA1 area, caused marked disruption or improvement in exploratory and cognitive behavior in non-obese mice fed with normal diet.
This study revealed that the consumption of diet containing high level of LCSFA, MCSFA, n-6 PUFA, MUFA, and TFA diets led to increases plasmic level of TC, HDL-C, and LDL-C, whereas diet high in n-3 PUFA induced reverse trend compared with the CON group in donor mice. The plasma TG were also increased in high LCSFA, n-3 PUFA, n-6 PUFA, MUFA, and TFA dietary groups. These results indicated that high-fat diets led to elevated blood lipids, and n-3 PUFA might be a protective role in the regulation of plasma cholesterol. Related research conducted by Micioni et al. and Opyd, indicated increases of plasma glucose, triglycerides and cholesterol, including HDL and LDL in obese rats fed hemp oil (4% diet) or hemp seeds (12% diet) [36, 41]. Lunn et al. demonstrated that the ap-propriate ratio of n-6/n-3 PUFA in diet is associated with improvement of blood lipid pro-file [42], which might be related to the different changes in plasmic TC, HDL-C, and LDL-C of obese mice induced by diet containing much n-3 PUFA in our study. Neverthe-less, FMT did not induce increases of plasma parameters in mice receiving “obese-type” gut microbiota compared with mice receiving “lean-type” gut microbiota. So the recipient mice did not replicate the obese phenotype of the donors from the results of current study.
Furthermore, our results demonstrated that the mice in most of high fat diet groups performed decreased cognitive functions with exception of mice consuming high n-3 PUFA diet. This may be relating to damages and structural breakage, and the fewer nerve cells in hippocampal CA1 area of mice in the LCSFA, MCSFA, n-6 PUFA, and TFA groups. Besides, the high LCSFA, n-6 PUFA, and TFA diet-induced impaired learning and memory ability was inherited in recipient mice by FMT, as showed in the novel object recognition test of their corresponding recipient groups. Similarly, the recipient mice in the + LCSFA, +n-6 PUFA, and + TFA groups performed reduced number of neurons and wid-ened intercellular spaces in hippocampal CA1 area. Interestingly, the n-3 PUFA group in donor mice and the CON-0 and CON-1 group in recipient mice showed significant differ-ences for exploring the two identical objects during the familiarity period of the novel ob-ject recognition tests. The possible reason was that some mice felt uncomfortable with new environment at the beginning, which made them spent a long time exploring the first contacting place. And during the test period, all mice have sufficiently adapted to the en-vironment. Overall, our results showed that high LCSFA, n-6 PUFA, and TFA diets dam-aged the structure of the hippocampal CA1 area by modulating gut microbiota, which in turn affected cognitive function. The control and high n-3 PUFA diets showed the opposite effect with them. These results are in consistent with associated epidemiological [43] and animal [8] studies, indicating that consumption of dietary fat is one of the most important risk factors in the development of cognitive dysfunction. Specially, excessive consumption of TFA and SFA was linked to obesity [44, 45] and neurologic conditions [8, 46]. Wu et al. revealed that high-fat diet-feeding could induce neuron cell injury, structural disorder, pro-inflammatory cytokine over-expression, and serotonergic system abnormality of the hippocampus in mice [47]. Previous studies have showed that PUFAs could improve the activity of brain cells, and enhance the learning and memory abilities in humans [48, 49]. De Mello et al. demonstrated that n-3 PUFA intake partially reversed the inflammatory, oxidative damage, and inhibition of mitochondrial respiratory chain complexes in the brain structures in obesity mice [50]. Similarly, Arendash et al. stated that high level of n-6 PUFA in cortex was related to cognitive decline in APP/PS1 transgenic mice [51]. However, some studies revealed negative association between intake of n-6 fatty acids and low cog-nitive performance [52, 53], which might be associated with anti-inflammatory and anti-oxidant effects of arachidonic acid. In addition, related studies found that low ratio of die-tary n-6/n-3 PUFA might decrease the risk of cognitive dysfunction [54, 55]. These differ-ences might be due to diverse research perspectives. More likely, independent discussion of n-6 PUFA intake might be associated with a lower risk of cognitive decline in animals or humans. The study of Jiang et al. showed that midlife dietary intake of n-6 PUFA was related to a lower risk of cognitive impairment in elderly Chinese participants [56]. Thus, above all, further exploration is required to study the beneficial effects on the cognitive abilities with appropriate doses of n-6 and n-3 PUFA, and the ratio between them. Several population studies reported that higher intakes of MUFA could improve cognition and prevent cognitive decline [56, 57], whereas others showed no significant associations for MUFA [58–60]. In our study, though high-MUFA diet-intake reduced the cognitive ability compared with basal-diet and high-n-3 PUFA diet-intake of mice in our study, high-MUFA diet-intake showed better cognitive behavior than high-MCSFA diet-intake. Both of our results and above literatures underscore different dietary fatty acids induced obesity could unequally affect in cognitive phenotype. While most of related publications indicted inflammatory responses and oxidative damage play key roles in high-fat di-et-induced cognitive impairment, the role of intestinal microbiota in it was emphasized in our study.
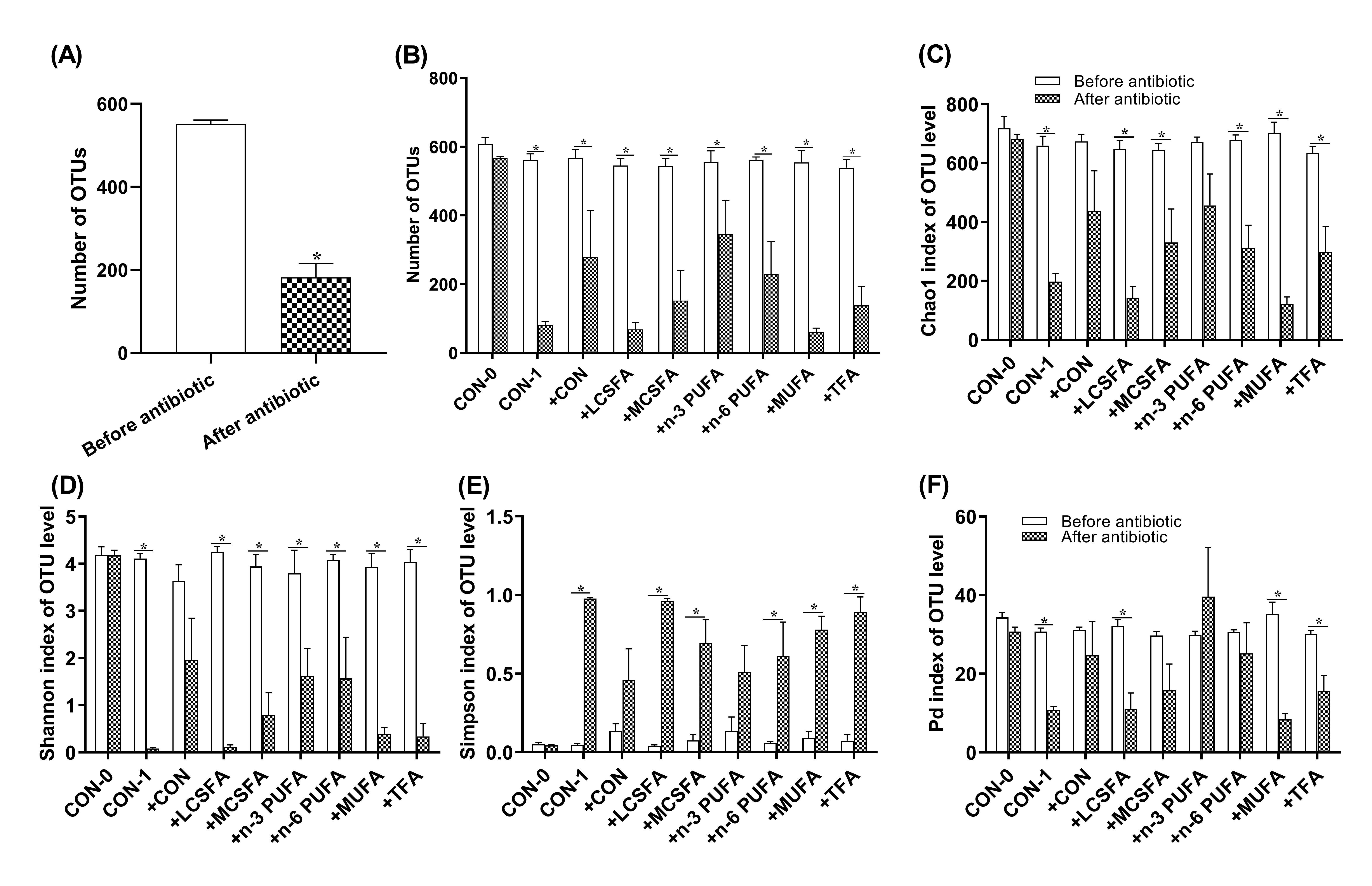
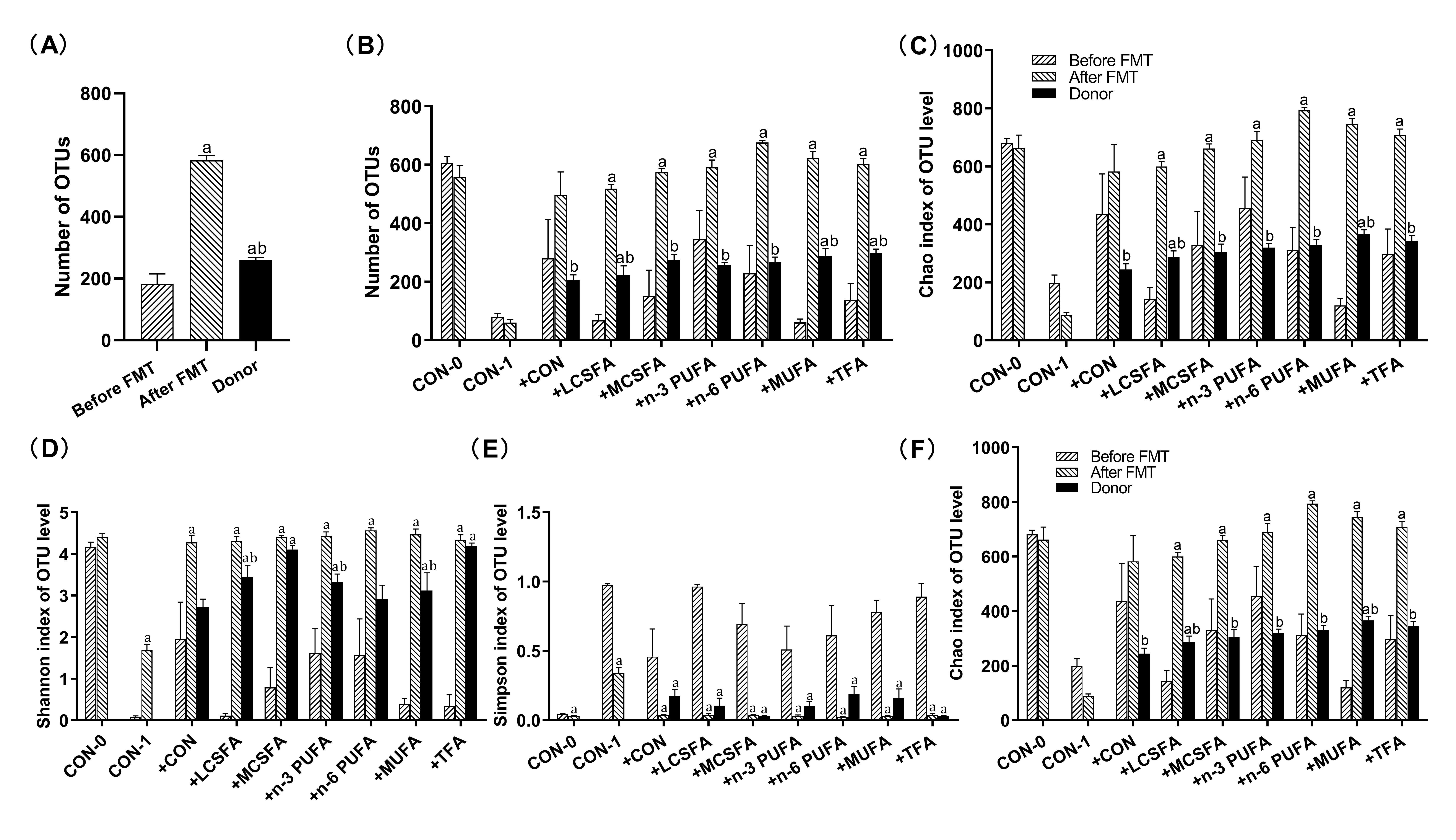
To explore the underlying mechanism of dietary fatty acids effecting hippocampal CA1 area, even cognition, the gut microbiota induced by multiple high-fat diets was transplanted to pseudo-germ-free mice (transplantation following antibiotics treatment). Related study demonstrated that FMT from aged donor mice induced impaired learning and memory ability, altered neurotransmission in hippocampus, and reduced the abun-dances of Lachnospiraceae, Faecalibaculum, Prevotellaceae and Ruminococcaceae in young re-cipients [27]. In our study, according to the alpha diversity, beta diversity, and the overall community difference analysis, we detected the intestinal microbiota of donor mice and in the three stages of recipient mice (before antibiotic treatment, after antibiotic treatment, and after FMT) and found similar alpha diversity all of the groups. Besides, the microbiota community diversity in MCSFA and TFA groups and the community richness in MCSFA, TFA, n-3 PUFA, n-6 PUFA and MUFA groups were higher than the CON group. These da-ta are contrary to that high-fat diets decreased microbial diversity or richness in most studies [61, 62]. Our results are similar to previous research reporting that high-fat diets can increase community diversity in mice [63, 64]. In recipient mice, the community rich-ness showed increases in the + n-6 PUFA, +MUFA, and + TFA groups after FMT, which was consistent with the conclusion of transplanted community keeping most of its original diversity [65].
Compared with the control, sixteen weeks of different high-fat-diets consumption induced extensive alteration in the intestinal microbial community structure at the phy-lum level in donor mice. The increased abundance of Firmicutes and decreased level of Bacteroidetes were observed in the six high fat feeding groups. These results are consistent with previous research revealing that high-fat diets induced increased Firmicutes and a decreased Bacteroidetes or a high Firmicutes/Bacteroidetes ratio [61, 63, 66]. These studies suggest that a higher ratio of Firmicutes/Bacteroidetes enables greater efficiency of gut mi-crobiota in extracting energy from the diets, which may be one of the reasons for obesity. In addition, increased Firmicutes are known as a risk factor in AD pathogenesis [67]. Hang et al. reported that FMT could ameliorate the cognitive impairment, and reverse the in-creased Firmicutes and the decreased Bacteroidetes of AD mice [68]. Tomova et al. reported that the fecal microbiome of autistic children showed a remarkable increase of the Firmic-utes/Bacteroidetes ratio [69]. In summary, the increased Firmicutes and the decreased Bac-teroidetes are not only associated with obesity, but also with cognitive dysfunction. Dinan et al. indicated that patients with irritable bowel syndrome showed an increase ratio of Firmicutes/Bacteroidetes and elevation of the amount of Lactobacillus spp, compared with healthy controls [70], which also suggests that these altered floras may play a role in au-tism through the "gut brain axis".
At the family level, the abundance of Muribaculaceae, which is mainly composed of genus norank_f__Muribaculaceae, showed decreases in the LCSFA, MCSFA, n-6 PUFA, and MUFA groups when compared with the CON and TFA groups of donor mice. In related studies, researchers found that the decreased Muribaculaceae was associated with reduced lipid and energy metabolism in AD mice [71] and consistent with clinical manifestations of AD patients [72]. In this research, there were higher abundances of Lachnospiraceae con-taining genus norank_f__Lachnospiraceae, in the LCSFA, MCSFA, and TFA groups than those in the CON and n-6 PUFA groups. Previous studies linked decreased Lachnospiraceae to neurodegenerative impairment [73], such as Parkinson’s disease [74]. Haskell-Ramsay et al. showed that nut supplementation exhibited a protective effect on cognitive function adults with the increased Lachnospiraceae in gut [75]. However, other studies suggested that different OTUs within the family Lachnospiraceae had different effects on place memory. Thus, we need to study it more deeply at the OUT level in the further research. We found mice in the MUFA group with a higher levels of family Erysipelotrichaceae and genus Faecalibaculum belonging to it than those in the LCSFA, MCSFA, and TFA groups. The enrichment of Erysipelotrichaceae showed relationships with cognitive dysfunction in AD rats [76]. Ruminococcaceae showed a higher abundance in the MCSFA and TFA groups than mice in CON, LCSFA, and MUFA groups, which was similar to previous study that indicated an increased expression of Ruminococcaceae in high-energy diets [77]. Neverthe-less, the family Ruminococcaceae associated with short-chain fatty acids production may beneficial to the structure of hippocampus [27]. The results of Bloemendaal et al. showed that probiotic supplementation in subjects with the increased Ruminococcaceae_UCG-003 could improve negative effects of stress on working memory [78]. In addition, our results showed increased abundance of family Lactobacillaceae and genus Lactobacillus in the n-6 PUFA group compared with those in the CON, MCSFA, n-3 PUFA, and TFA groups. Many researches demonstrated that Lactobacillus, as a probiotic, enhanced memory and changed regional brain metabolites in humans and animals [79, 80]. These studies are contrary to our results, and it may be necessary to further study the relationship between them and the metabolic reactions of different fatty acids. In summary, increased richness of Firmicu-tes phylum, families of Lachnospiraceae, Erysipelotrichaceae, Ruminococcaceae, and Lactobacil-laceae, genera of norank_f__Lachnospiraceae, Faecalibaculum and Lactobacillus, and decreased level of Bacteroidetes phylum, Muribaculaceae family and norank_f__Muribaculaceae genus in gut microbiota were found in high-fat diet-induced obesity mice with low cognitive per-formance in current study. Further studies are needed to investigate the roles of these gut microbiota in their effects of high-fat diet on cognitive function.
After transplantation of donor gut microbiota, the relative abundances of phyla Ver-rucomicrobia, Proteobacteria, and family Akkermansiaceae showed significant increases, whereas families Muribaculaceae, Prevotellaceae, Ruminococcaceae, and Rikenellaceae, genera norank_f__Muribaculaceae, Prevotellaceae_UCG-001 and alistipes showed marked decreases in the CON-1 group when comparing with other groups. Related reports indicated that the increased Patescibacteria in patients with AD or mild cognitive impairment was correlated with Clinical Dementia Rating [81]. The phylum Patescibacteria is associated with cyto-kines from “gut-brain axis” including brain-derived neurotrophic factor, serotonin, and others [82]. Contrary to the donor mice, the abundance of Muribaculaceae and genus nor-ank_f__Muribaculaceae in the + n-3 PUFA group was decreased comparing with the CON-0, +MCSFA, +n-6 PUFA and + TFA groups. In one previous study, the Chitooligosaccharides has been verified to promote neuronal regeneration in hippocampus by decreasing the relative level of Muribaculaceae [83]. This reverse change might be caused by antibiotic treatment and/or FMT. The abundance of Ruminococcaceae, was decreased in the + MCSFA group when compared to CON-0 group, which was similar to the previous study finding the decreased Ruminococcaceae in young recipient mice from old donors by FMT [27]. The abundance of Rikenellaceae in the mice of + LCSFA, +n-3 PUFA, +n-6 PUFA, +MUFA and + TFA groups, meanwhile the genus alistipes which is assigned to Rikenellaceae in + LCSFA, +n-6 PUFA and + MUFA groups, showed increases compared to the + CON group. The finding of Ren et al. indicated a higher abundance of Rikenellaceae in Parkinson’s disease patients with mild cognitive impairment compared with healthy subjects [84]. Lee et al. indicated that Rikenellaceae was correlated with cognitive function and blood and fecal LPS levels in mice [85] which showed the changes in intestinal flora might change cogni-tive function by stimulating inflammatory response.
Our results suggested that this binary distinction did not sufficiently reflect the com-plex influence of diet-induced gut microbiome on cognition, as reported in the previous study of Bruce-Keller and his colleagues [1]. These deviations could be induced by the dif-ferent sequencing depth between fecal samples collected from donor mice and recipient mice in different time points. Moreover, the CON-0 and CON-1 group could express the effects of repeated gavage and antibiotic treatment respectively. Other groups were treated equally to minimize the potential artifacts. Appropriate animal models are indispensable in exploring the association between the host and the intestinal flora [86]. However, germfree animals have notably underdeveloped immune systems, decreased cardiac output [87] as well as delay gastric emptying and gastrointestinal transit [88]. Therefore, antibiotic-treated mice are more suitable than germ-free mice for the current study because it can resemble the human condition.
According to our results, the similar behavioral phenotype between donor mice and recipient mice combined with the transplantation of diet-induced intestinal microbiota supported the concept of a “microbiome-gut-brain axis”. Khan et al. reported that mice were exposed to the increase of social defeat stress, which caused the higher attenuation of Lactobacillus [89]. Moreover, some studies indicated that probiotic intake is linked to posi-tive changes in behavior and mood [90]. “Gut microbiota, gut, and brain could interact via the central nervous system, either through the enteric nervous system, via spinal and va-gal nerves, or via blood circulation” [91, 92]. But the detailed mechanism of the gut micro-biota’s effects on cognitive function in mice needs to be further studied.
Taken together, high LCSFA, MCSFA, n-6 PUFA, MUFA, and TFA diets induced obe-sity, increased plasmic cholesterol, and the reverse regulation of high n-3 PUFA diet were evident in the donor mice. However, these obesity phenotypes were not induced in recipi-ent mice by FMT. The high LCSFA, n-6 PUFA, and TFA diet-induced impaired spatial learning and memory ability in donor mice was inherited in recipient mice by FMT. On the contrary, the high n-3 PUFA diet might be a protective factor of cognitive function. High LCSFA, MCSFA, n-6 PUFA, and TFA diets induced structural breakage and reduced nerve cells in hippocampal CA1 area in donor mice, which was also showed in corresponding groups (+ LCSFA, +MCSFA, +n-6 PUFA, and + TFA groups) of recipients. In addition, increased phylum Firmicutes, family Erysipelotrichaceae and genus Faecal-ibaculum, and decreased phylum Bacteroidetes, family Muribaculaceae and genus nor-ank_f__Muribaculaceae in gut microbiota of donor mice might be positively correlated with low cognitive performance in high-fat diet-induced obesity donor mice. In the same way, increased phylum Patescibacteria, family Rikenellaceae and genus alistipes, and decreased family Ruminococcaceae in recipient mice might be associated with cognitive impairment.
Our study has strong limitations by using mice as study subjects and the different sequencing depth may lead to unexpected results for microbiota analysis. The results in mice studies do not always extend into human studies and relevant population studies are very necessary. Thus, in-depth study of which bacteria in the intestinal flora play a role in the process of FMT causing similar phenotypes between recipient mice and donor mice should be the focus of future research. Besides, this study suggested that the therapeutic manipulation of the microbiome in humans should be deeply studied and utilized in neuropsychiatric diseases.
{kind=link}
{kind=link}