The current study was conducted to determine how bacterial inoculation with a multifunctional plant growth promoting M. oryzae CBMB20 affects the expression of proteins in rice plants, IR29 and FL478, and promotes plant growth and development during the rapid changes occurring during the early seedling development. We initially observed the endophytic colonization of CBMB20 in IR29 and FL478. To establish inherent phenotypic proteomic differences between IR29 and FL478, IR29-NI and FL478-NI were compared. To assess microbe-mediated proteomic changes due to CBMB20 inoculation, treatment comparison between IR29-NI vs. IR29-I and FL478-NI vs. FL478-I were analyzed. Additional information could also be derived in the inoculated plants by comparing IR29-I vs. FL478-I.
Inherent Phenotypic Similarities And Differences Between Ir29 And Fl478 Expressed Through Their Proteomes
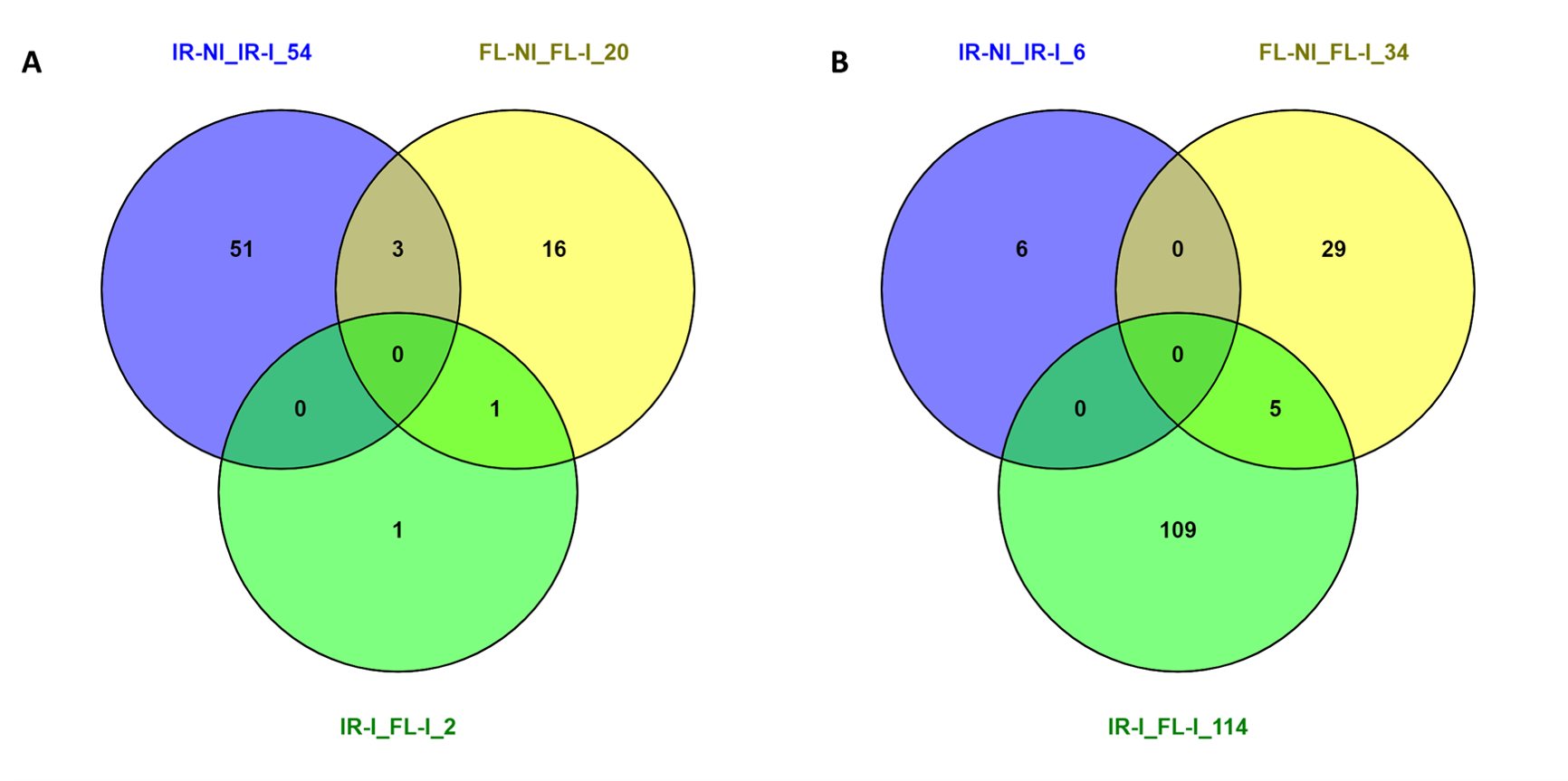
The cultivars IR29 and FL478 are genotypically closely related rice cultivars where FL478 traces its parental lineage to IR29 and Pokkali of which IR29 is the high yielding salt-sensitive parent and Pokkali is the salt-tolerant low yielding parent (Walitang et al., 2019). The present study validates the close relatedness of the two rice genotypes as almost 88% of their phenotypically expressed proteins during their early seedling development are exactly similar. However, the level of expression of these proteins potentially differs between the two rice cultivars as we observed 28 DAPs in salt tolerant FL478 compared to 52 DAPs in the sensitive rice variety IR29. The remaining differences in expressed proteins may further lead to distinct differences in IR29 and FL478. The differences in certain biological functions and processes observed in this study corroborated the transcriptional and translational differences between the parent genotypes observed in an earlier study (Li et al., 2018).
Overall, a majority of the DAPs associated to FL478 and IR29 are related to metabolism, protein synthesis, stress and defense, carbohydrate and energy metabolism, membrane and transport. However, inherent differences between the two rice genotypes become apparent especially under salt stress conditions in which proteomes and transcripts points to variations in salt-stress adaptive defenses between the two rice cultivars.
Previous comparative studies under salt stress between the two rice genotypes showed that FL478 show greater abundance and upregulation of genes and proteins associated to superoxide dismutase, peroxidase genes, fiber protein, and inorganic pyrophosphatase with concurrent higher relative growth rate and lower Na+/K+ in the roots (Hosseini et al., 2015; Senadheera et al., 2012). Differences in biological process as well as molecular function were also observed between IR29 and FL478 through their transcriptional profile (Mirdar Mansuri et al., 2019) which corroborates the observed differences in terms of proteomics in the present study even under normal conditions. The observed differences between the two rice genotypes under normal growing conditions could be mainly attributed to the differences in gene expression profiles, both transcriptome and translatome, as seen in the comparison between IR29 and FL478’s paternal parent, Pokkali, showing distinct differences between the two even without salt stress (Li et al., 2018). Under normal conditions, Li et al. (2018) observed that the gene ontology terms overrepresented in Pokkali include regulation of metabolism, gene transcription, cellular carbohydrate metabolism, and DNA conformation while photosynthesis, response to stress, cell wall macromolecule metabolism and organization, transport and aminoglycan metabolism were observed for IR29. This pattern is quite similar to the proteomes of IR29 and FL478 especially after inoculation with CBMB20.
Changes in the functional proteomes gene ontology terms in IR29 and FL478 induced by inoculation of the multi-faceted Methylobacterium oryzae CBMB20
Methylobacterium oryzae CBMB20 has been well documented in enhancing plant growth under normal (Lee et al., 2006; Ryu et al., 2006; Madhaiyan et al., 2010) and stress conditions (Indiragandhi et al., 2008; Yim et al., 2013; Yim et al., 2014; Chanratana et al., 2018; Chatterjee et al., 2019). Common features of M. oryzae CBMB20 plant growth promotion usually include enhanced germination, germination rate, and seedling vigor index together with increased shoot and root length and increased biomass. However, there are potential differences in the responses across crop plants due to inoculation of CBMB20 (Ryu et al., 2006; Madhaiyan et al., 2010). In the current study, distinct differences in cultivar responses even at proteome level could be observed between IR29 and FL478. Interestingly, inoculation with CBMB20 expanded the gene ontology terms of the different proteins in both IR29 and FL478. The proteomes of IR29 and FL478 presented a much more diversified gene ontology terms upon inoculation with CBMB20. This supports the idea that inoculation with CBMB20 has extended the phenotype of the rice plants (Dawkins, 1999; Hardoim et al., 2015) mediated by the microbe-responsive proteomes.
Comparing the functional categories of the proteins associated to the CBMB20 inoculated IR29 and FL478 shows that there are more proteins related to photosynthesis, metabolism and protein degradation in IR29 while more proteins are associated to metabolism, stress and defense, cell structure, differentiation and fate, and protein folding and transport were observed in FL478. Roy Choudhury et al. (2022) showed that under normal growing conditions, inoculation with CBMB20 of a rice japonica cultivar increased the chlorophyll contents and carotenoid content of the leaves. In the present study, a protein directly involved with pigment synthesis (NADPH-protochlorophyllide oxidoreductase – A2XZ01) and chlorophyll-carotenoid formation was upregulated in IR29. Other photosynthesis-related proteins detected are mainly involved in the integrity and function of the chloroplast. The upregulated proteomes of IR29 and FL478 due to CBMB20 seem to reflect the enriched functional categories of transcriptomes and translatomes in IR29 and Pokkali (Li et al., 2019) and may point to a notion that CBMB20 inoculation reinforces these inherent differences in the two genotypes. Earlier, metabolomic and transcriptomic changes in Arabidobsis inoculated by Pseudomonas fluorescens SS101 has been established to be a strain-specific response (van de Mortel et al., 2012). However, our study showed that there are also plant genotype-specific proteomic responses occurring due to inoculation such as enrichment of photosynthesis-related proteins in IR29 and proteins related to cell differentiation in FL478. Although CBMB20 is an efficient plant growth promoter in multiple crop plants, the observed plant genotype-specific responses could explain differences in the effectiveness of bioinoculants and the need to investigate compatibility of crop and microbial inoculants such as those observed under field conditions (Sanjenbam et al., 2022).
Common microbe-responsive proteins modulated by Methylobacterium oryzae CBMB20
Although there is a difference in the overall response of IR29 and FL478 to CBMB20 inoculation, there are similar proteins upregulated indicating similar modes of microbe-mediated proteomic changes in both rice genotypes. There were three proteins both upregulated in IR29 and FL478 upon CBMB20 inoculation viz peptidyl-prolyl cis-trans isomerase (A2WJU9), thiamine thiazole synthase, chloroplastic (A2YM28), and alanine—tRNA ligase, chloroplastic / mitochondrial precursor (B8B4H5).
The enzyme peptidyl-prolyl cis-trans isomerase catalyze the isomerization between the cis and trans form of peptide bonds (Lin et al., 2019) particularly certain proline residues involved in protein folding (Blecher et al., 1996) and cellular protein modification process (GO: 006464). This enzyme is also regulated by various environmental stresses and could act as a molecular timer in diverse plant physiological and pathological processes (Blecher et al. 1996). Thiamine thiazole synthase is essential for thiamine biosynthesis especially targeted to chloroplasts influencing chloroplast development (Feng et al., 2019). However, the thiamine thiazole synthase detected in IR29 and FL478 is also related to response to stimulus (GO: 0050896) and different metabolic processes (GO: 0006727 – aromatic compound, GO: 0006777 – vitamin, GO: 0006790 – sulfur compound, GO: 0006807 – nitrogen compound, and GO: 0046483 – heterocycle metabolic process). In addition, based on gene ontology, alanine—tRNA ligase chloroplastic/mitochontrial precursor (B8B4H5), which attaches alanine to tRNA, is associated with translation (GO: 0006412), cellular amino acid metabolic process (GO: 0016070), and RNA metabolic process (GO: 0016070). The upregulation or enrichment of exactly similar proteins or proteomes with analogous functions in the two rice genotypes is potentially due to the same responses brought about by CBMB20 producing metabolites and proteins involved with plant growth promotion including hormones (Lee et al., 2006; Ryu et al., 2006; Madhaiyan et al., 2010) and improvement of plant nutrient contents (Anandham et al., 2007; Madhaiyan et al., 2010; Kwak et al., 2014).
Methylobacterium oryzae CBMB20 has been also observed to induce defense responses and was able to increase resistance to subsequent biotic stresses indirectly promoting growth through plant health (Indiraghandi et al., 2008; Yim et al., 2013; Yim et al., 2014). Colonization of non-native bacterial endophytes usually results in the activation of the plant’s defense responses and the colonizer has to either evade or modulate these responses in order to beneficially coexist with their host (Pieterse et al., 2014; Balmer et al., 2015). Several defense-related proteins have been modulated after rice inoculation with CBMB20. In FL478, thioredoxin H-type (A2YIW7) and glutathione transferase (A2Z9L8) were repressed similar to the thioredoxin domain-containing protein (A2YL83) observed in IR29. These proteins are important for regulating redox reactions and redox-mediated defense signaling (Yoo et al., 2011; Hosseini et al., 2015; Chi et al., 2019). These stress responsive proteins are potentially regulated indirectly though reduction of stress ethylene (Indiraghandi et al., 2008; Yim et al., 2013; Yim et al., 2014). These results show that CBMB20 has the capacity to modulate defense responses in rice and different from those observed in pathogenic bacteria (Pieterse et al., 2014; Balmer et al., 2015) where their presence lead to hypersensitive response or induction of systemic acquired resistance aimed at eliminating the invading pathogens. The modulation of stress responses by CBMB20 contributes to the indirect mechanism of plant growth promotion as CBMB20 inoculation potentially results to tolerance enhancement when rice plant is exposed to biotic and abiotic stress.
{kind=link}