According to the data of World Health Organization (WHO), hepatocellular carcinoma is the most common primary liver tumor in adults and the sixth most common cancer in the world, and the mortality rate is the third in the world, which is one of the major malignant tumors threatening human health[1, 14, 15]. The main reason for this situation is that viruses, chemicals, congenital or acquired metabolic diseases are the causes of HCC, and it has the characteristics of great harm to the human body and a wide range of body damage. At the same time, there is a lack of clinical biomarkers that can be used to diagnose HCC with high sensitivity and high specificity[16]. Combined with the concealed pathogenesis characteristics of HCC, most patients have entered the middle and advanced stage when diagnosed. In recent decades, the treatment of HCC has made some achievements by the development of radiotherapy, chemotherapy, targeted immunotherapy, surgical treatment and clinical practice. Thanks to new therapeutic methods, such as local ablation, mid-term chemoembolization, advanced systemic therapy, targeted therapies, the establishment, development and maturity of liver transplantation technology, and the proposal of precision medicine, the treatment of HCC has gained certain benefits. However, there is still a bottleneck in the treatment of HCC. Despite the use of various therapies to treat HCC, the prognosis of HCC remains poor due to the high frequency of metastasis and recurrence, with a poor five-year survival rate[17–19]. Therefore, it is necessary to find biomarkers that can diagnose HCC as early as possible and to study more molecular pathways and possible resistance mechanisms. That is still the goal of HCC treatment. In recent years, the use of integrated multiple omics analysis has made significant progress in gaining insight into the molecular markers involved in the initiation and progression of HCC and has contributed to a comprehensive mapping of key signaling pathways and aberrant molecular events in HCC. In this context, targeted therapy has become the historical stage for the treatment of advanced HCC. For the vast majority of patients with advanced HCC who cannot be operated on or have extensive postoperative recurrence, targeted therapy is an extremely important treatment. The occurrence and progression of HCC is always associated with the dysregulation of various cellular mechanisms (such as proliferation, survival, angiogenesis, etc.) caused by viral infections, alcoholism, metabolic diseases, and exposure to carcinogens[20]. The targeted drugs currently used to treat HCC can inhibit these cellular processes by targeting multiple signaling pathways. Among the available therapies, sorafenib, lenvatinib, the combination of atezolizumab and bevacizumab are the first-line systemic treatments for HCC, regorafenib, ramucirumab, and cabozantinib are second-line systemic treatments[21]. In 2008, sorafenib became the first drug approved for advanced HCC. Subsequently, lenvatinib was proved to be no inferior to sorafenib in phase III multicenter trials[22], and in recent years, lenvatinib has become the most commonly used targeted therapy drug of choice for the treatment of advanced HCC due to its good tolerance. However, the high drug resistance of HCC is the main problem limiting its overall efficacy. Currently, targeted drugs used in the systematic treatment of HCC are a class of multi-target kinase inhibitors, which can act by inhibiting Vascular Endothelial Growth Factor Receptor (VEGFR), Raf/mitogen-activated protein (MAP) kinase/extracellular signal-regulated kinase (ERK) (Raf/MEK/ERK), Platelet Derived Growth Factor Receptor (PDGFR) and other pathways[23]. However, from the perspective of clinical practice, targeted drug therapy effect is not ideal, only some patients can benefit from it, and some patients have more or less primary drug resistance or secondary drug resistance. According to statistics, the vast majority of patients who have received effective sorafenib treatment will develop different degrees of drug resistance within 6 months[24]. At present, the mechanism of targeted drug resistance in HCC is not clear. In recent years, studies on the resistance of targeted drugs in HCC mainly focus on: (1) When drug resistance occurs, the increase of stemness of HCC cells is considered to be one of the important reasons for drug resistance[5]; (2) Counteracting the effects of targeted drugs on inhibiting RAF, VEGFR and other pathways by activating AKT, mTOR and other pathways through bypass[25]; (3) The mechanism of autophagy and Epithelial Mesenchymal Transformation (EMT) may also be the cause of drug resistance in HCC[10]; (4) The expansion of Epidermal Growth Factor Receptor (EGFR) is one of the main causes of lenvatinib resistance in HCC[6]; (5) In addition, the research has found that inhibiting the expression of cell cycle mcl-1/cyclin E1 can promote drug resistance of HCC, and introducing drugs that inhibit cyclin-dependent Protein Kinase (CDK) can improve the efficacy of targeted drugs[26]. These results suggest that the resistance mechanism of targeted drugs is extremely complex in HCC, and there must be other molecular mechanisms that are more complex and variable. Therefore, it is urgent to clarify the new mechanism of targeted drug resistance in clinical practice, and explore new molecular markers and molecular mechanisms, which will have great clinical significance for improving the overall efficacy of HCC.
It is well known that tumors involve extensive variation in genomics, transcriptomics and proteomics. Most previous transcriptomics studies focused on the dynamic changes of linear transcripts in the process of tumor genesis and development, while ignoring the role of circRNA. However, in recent years, circRNA has been frequently reported to play a non-negligible coordinating role in multiple biological processes of tumor development. CircRNAs are a class of single-stranded molecules with tissue and development-specific expression patterns, and recent studies have shown that the biological characteristics of circRNAs mainly focus on the following four aspects[27, 28]: (1) Stability, because its unique circular structure makes circRNAs have relatively strong resistance to ribonuclease, so it is more stable than linear RNA; (2) High abundance, thousands of different circRNAs have been identified in eukaryotes, with extremely rich species and number; (3) Conservation, the expression patterns of circRNAs show evolutionary conservation across different species; (4) Specificity, circRNAs usually show specific expression in different tissues or developmental stages. These remarkable features of circRNAs have stimulated the drive to unravel the mystery of their biological function. Interestingly, a growing number of studies have also shown that circRNAs play a very important regulatory role in tumor drug resistance[29]. More importantly, many studies have found that circRNA has a significant effect on targeted drug resistance of HCC. One study had found the circRNA-SORE (an increased expression circRNA in sorafenib resistant HCC cells), which can bind to the major oncogenic protein YBX1 in the cytoplasm and block the nuclear interaction between YBX1 and E3 ubiquitin ligase PRP19, thus blocking the PRP19-mediated degradation of YBX1, and thereby enhancing sorafenib resistance in hepatocellular carcinoma[8]. In another study by the researchers, it was found that after m6A modification promoted circRNA-SORE expression, circRNA-SORE could separate miR-103a-2-5p and miR-660-3p by acting as miRNA sponge, thus competitively activating the Wnt/β-catenin pathway and inducing sorafenib resistance[9]. In addition, research has found that the high expression of circUHRF1 in HCC can inhibit the secretion of IFN-γ and TNF-α by NK-cell-derived, and inhibit the function of NK cells by degrading miR-449c-5p and upregulating the expression of TIM-3, thus leading to the resistance of HCC patients to anti-PD1 treatment[30]. Other studies have found that circMET is overexpressed in HCC, which can drive the immunosuppression and anti-PD1 therapeutic resistance of HCC through Snail/DPP4/CXCL10 axis[31]. Besides, circFN1 has also been reported to mediate sorafenib resistance in HCC by adsorbing miR-1205 and regulating the function of transcription factor E2F1[32]. These results fully demonstrate that circRNA plays a key regulatory role in the occurrence and development, diagnosis and prognosis, mediating drug resistance and other processes of HCC, and it is necessary to explore the upstream and downstream regulatory molecular mechanisms. Therefore, our study further investigated the effect and mechanism of circRNA related to HCC on the progress and drug resistance of HCC.
Autophagy is an evolutionarily conserved endogenous catabolic activity that regulates lysosomal degradation of cell components and damaged organelles. Autophagy supports cell survival and maintains homeostasis in response to different forms of stress, such as hypoxia or lack of nutrients and energy[33]. In addition, autophagy can promote the invasion and migration of HCC cells[34, 35]. However, whether and how autophagy contributes to cancer progression remains controversial. It has been suggested that by making residual or metastatic cancer cells tolerant to cytotoxic stress, autophagy makes them drug-resistant[36]. Based on the cytoprotective properties of autophagy in cancer cells, most of the current studies on autophagy focus on exploring the value of autophagy targeted therapy[37]. Due to the complex role of autophagy in HCC (homeostasis, signaling, and potential prognostic and therapeutic value)[38], it is an extremely important and unmet need to use this point to develop autophagy-related methods as a therapeutic strategy for HCC and improve the therapeutic effect. Recently, many studies have shown that circRNA is also closely related to tumor autophagy and tumor drug resistance. Guo et al. found that hsa_circ_0023404 could significantly affect the expression of autophagy-related genes Beclin1 and p62 in cervical cancer cells, and enhance cervical cancer metastasis and chemotherapy resistance through VEGFA and autophagic signaling via sponge miR-5047[39]. Moreover, autophagy may also be an important mechanism of targeted drug resistance in HCC[10]. These findings fully demonstrate that autophagy mechanism is closely related to circrNA-induced tumor progression and drug resistance. However, up to now, whether autophagy-related circRNA plays a role in the progression and the resistance of targeted drugs of HCC and its related mechanism are rarely mentioned in the literature. Therefore, in this study, we first used Limma package to perform differential expression analysis and pathway enrichment analysis on circRNA related data in the database, and obtained differentially expressed autophagy-related circRNAs from control sequences. We then identified the key circRNA (hsa_circ_0009792) by multiple identification and validation experiments.
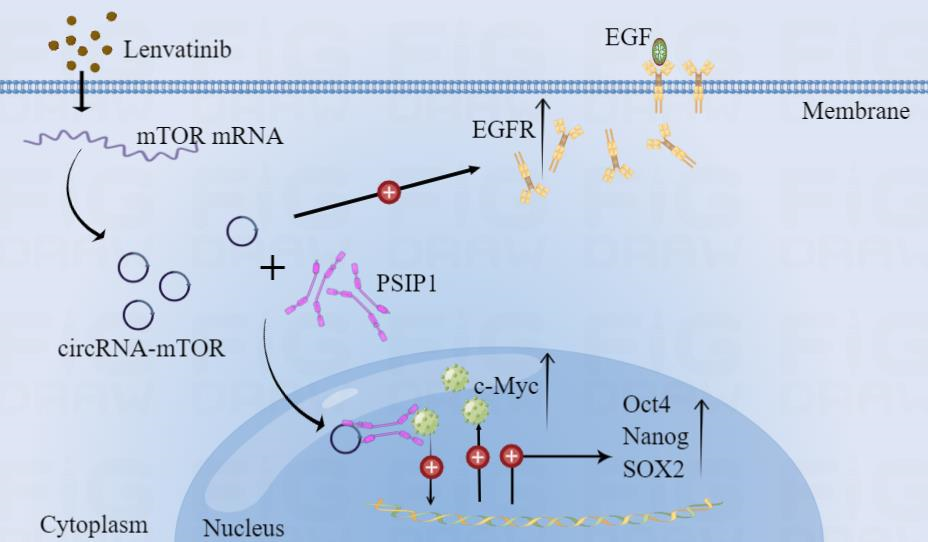
By querying circBase database and UCSC database, we found that hsa_circ_0009792 was formed by cyclization of exons 48 and 49 of mTOR gene. Since the mother gene of hsa_circ_0009792 is mTOR, hsa_circ_0009792 in this study is also called circRNA-mTOR. Then, in order to further confirm that circRNA-mTOR is an important circRNA with clinical significance, we analyzed the relationship between hsa_circ_0009792 and various clinical indicators. Through our analysis of relevant clinical samples and data, we found that circRNA-mTOR has potential important diagnostic value and clinical significance. Although there are various forms of circRNA formation, according to the source of circRNA, circRNA can usually be divided into: exonic circRNA (ecRNA), intronic circRNA (ciRNA) and exon-intron circRNA (EIciRNA), and the difference of intracellular localization determines the difference of its function to a certain extent[11, 12, 27]. In this study, we used nucleoplasmic separation and FISH experiment to explore the localization of circRNA-mTOR in HCC cells. The experimental results showed that circRNA-mTOR exists in both the nucleus and the cytoplasm, and the proportion of circRNA-mTOR is slightly higher in the nucleus. It is helpful for us to design and synthesize the overexpression or interference system, and also has extremely important guiding significance for us to further explore the mechanism. To investigate the relationship between circRNA and HCC, it is first and foremost important to explore whether the circRNA to be studied have any effect on hepatocellular carcinoma. Previous studies have shown that the effects of circRNA on HCC are multifaceted. In our study, we proved that circRNA-mTOR can promote the growth and proliferation, clone formation, invasion and metastasis, and reduce the apoptosis of hepatocellular carcinoma by cell function assay and in vivo tumorigenesis assay. In other words, circRNA-mTOR could significantly promote the progression of HCC. As mentioned above, circRNAs are closely related to the drug resistance of HCC, so the effect of circRNA-mTOR on the drug resistance of HCC need to be explored. The IC50 assay was used to detect the changes of the IC50 value of lenvatinib in different HCC cell lines. The results showed that the IC50 value of lenvatinib increased to different degrees after overexpression of circRNA-mTOR, while the IC50 value of lenvatinib decreased to varying degrees after the expression of circRNA-mTOR was interfered. This indicated that circRNA-mTOR has a significant promoting effect on lenvatinib resistance in HCC. In order to further explore, we treated HCC cells with lenvatinib, and confirmed that circRNA-mTOR could promote the growth and proliferation, clone formation, and reduce the apoptosis of HCC cells through cell function experiments. In other words, circRNA-mTOR could promote lenvatinib resistance in HCC.
Previous studies have shown that increased tumor stemness is considered to be one of the main reasons for the development of drug resistance in HCC[5]. A group of stem/progenitor cells called tumor-initiating cells or cancer stem cells (CSC) has been found to promote changes in tissue hierarchy and enhance the generation of heterogeneity[40]. Different from normal tumor cells, these tumor stem cells show a strong ability to self-renew, differentiate and form new tumors, which in turn reinforces the resistance to conventional therapy and the high rate of tumor recurrence[41]. Similar to stem cells in other tissues of the body, tumor stem cells depend on the activation of highly conserved stem signaling pathways to maintain self-renewal and regeneration[42]. Notably, aberrant activation of these pathways in stem cells may cause uncontrolled cell proliferation and aberrant differentiation leading to tumorigenesis, while their reactivation can induce tumor reprogramming leading to the emergence of CSC phenotypes. These pathways also contribute to the occurrence and recurrence of HCC. In recent years, a lot of research has been done on HCC related CSCs, but the mechanism of maintaining the self-renewal of these CSCs is still difficult to understand[43]. With the deepening of research, a large number of studies have shown that circRNA and other small molecular RNA are closely related to tumor stemness of HCC. In the study of Gu et al., CircIPO11 was found to be highly expressed in HCC tumor tissues and liver CSCs, CircIPO11 is required for the self-renewal maintenance of liver CSC to initiate HCC development, and circIPO11 can recruit TOP1 to the GLI1 promoter to trigger its transcription and active Hedgehog signaling[44]. Another study has shown that circZKSCAN1 deletion confers a variety of malignant properties, including cancer stem, and is closely related to overall and recurrence-free survival of HCC[45]. CD90 can promote pellet formation ability of HCC cells through hsa_circ_0067531, that is, improve tumor stemness of HCC[46]. Circ_0007059 is down-regulated in HCC tissues and cells, which can enhance cell apoptosis and inhibit the proliferation, migration, invasion and stem cell-like properties of HCC cells[47]. At present, it is generally accepted that the sphere formation assay (the number and size of sphere) is the gold standard to measure the stemness level of tumor cells. Based on this, the result of our sphere formation assay showed that circRNA-mTOR could improve the sphere formation ability (size and efficiency of sphere formation) of HCC cells treated with lenvatinib, which indicated that circRNA-mTOR could promote lenvatinib resistance by improving stemness level of HCC cells to a certain extent.
In one study, circ_MMP2 (has_circ_0039411) was found to be delivered to other cells via exosomes derived from HCC cells and was associated with reduced overall survival in HCC patients, and it can up-regulate its parent gene MMP2 through the sponge effect of miR-136-5p and promote HCC metastasis[48]. In another study, CircC16orf62, as a molecular sponge of miR-138-5p and a competitive endogenous RNA of PTK2, promoted the activation of autophagy pathway to promote the progression of HCC[49]. In addition, circ_0007059 can inhibit the stem cell-like properties of HCC and inhibit the autophagy pathway by positively regulating the expression of phosphatase and tensin homolog (PTEN)[47]. These studies suggested that circRNAs could exert their biological effects by affecting their parent genes or autophagy in HCC. So, does the circRNA-mTOR we studied also affect its parent gene? And since circRNA-mTOR was identified through the autophagy pathway, and its parent gene mTOR was also a key protein in the autophagy pathway, we first explored the influence of circRNA-mTOR on its parent gene mTOR and autophagy pathway. However, it was interesting that circRNA-mTOR had no obvious effect on mTOR and autophagy pathway, which required us to find its mechanism additionally.
According to previous studies, we found that circRNA can play its biological functions at multiple levels: (1) Acting as miRNA inhibitors to regulate gene expression at the post-transcriptional level ("sponge" effect)[7, 50, 51]; (2) Regulation of gene transcription and RNA splicing[12, 52, 53]; (3) As a protein bait to regulate protein function[54]; (4) Encoding functional peptide[55]; (5) Other confirmed functions[56–58]. However, it is known that miRNAs target specific mRNAs through the principle of base complementary pairing, and the "sponge" function of circRNAs is to exert its Competing Endogenous RNAs (ceRNAs) by competitive binding to miRNAs, and that are then regulated at the post-transcriptional level. This process generally occurs in the cytoplasm, but we had shown previously that the cellular localization of circRNA-mTOR existed in both the nucleus and the cytoplasm, and the localization was slightly more in the nucleus, so we inferred that circRNA-mTOR did not play its biological function through "sponge" miRNA. In addition, generally speaking, circRNAs need to have two basic conditions to function as coding peptides, one is an open reading frame (ORF), the other is an internal ribosome entry site (IRES). Based on this we through database ORF finder (https://www.ncbi.nlm.nih.gov/orffinder) to query the circRNA-mTOR presence of ORF, through database IRESite (http://iresite.org/IRESite_web.php) to query the presence of IRES. By comparing the matching results and cell localization results, we found that circRNA-mTOR had a low possibility of translation function. Else, other functions of circRNAs generally refer to fusion-circRNA. The circRNA-mTOR we studied was not a fusion-circRNA, so the possibility of it playing other functions was also very low. Therefore, we suggested that circRNA-mTOR may act as a protein decoy to regulate protein function or play a role in regulating transcription/differential cleavage. Numerous previous studies have shown that the interaction between RNA-binding protein (RBP) and circRNAs is important for the regulation of tumor progression[13]. In HCC, it has been reported that circRNAs interact with RBP to produce biological effects. The study had found that circRHOT1 could recruit TIP60 to the promoter of NR2F6 and initiate NR2F6 transcription to promote the progression of HCC[59]. CircDLC1 could bind to the RBP (HuR), thereby reducing the interaction between HuR and MMP1, thereby inhibiting MMP1 expression and ultimately contributing to the inhibition of HCC progression[60]. CircETFA can promote the progression of HCC by recruiting the RNA-binding protein (EIF4A3) to prolong the half-life of CCL5 mRNA[61]. Downregulation of hsa_circ_0074854 can suppress the migration and invasion of HCC by interacting with RNA-binding protein (HuR) and inhibiting exosome-mediated M2 polarization of macrophages[62]. ESR1-activated circ_0004018 inhibits angiogenesis in HCC by binding to RNA-binding protein (FUS) and stabilizing TIMP2 expression[63]. RNA-binding protein RBM3 promotes HCC cell proliferation by interacting with circular RNA SCD-circRNA 2[64]. CircCPSF6 attenuates the binding of PCBP2 to YAP1 mRNA through competitive binding, thereby maintaining the stability of YAP1 and promoting the malignant progression of HCC[65]. Therefore, in order to explore the RBP binding to circRNA-mTOR, we conducted RNA pulldown and LC-MS experiment. At the same time, based on tumor stemness enhancement, combined with LC-MS results, functional analysis and protein classification analysis results, we screened PSIP1 as the key RBP protein to be studied from the differential proteins through bioinformatics analysis, previous literature analysis, and main function analysis of RBP. By querying Gene Cards database (https://www.genecards.org), we found PSIP1 (PC4 and SFRS1 Interacting Protein 1) is a protein-coding gene, also known as LEDGF/p75, which encodes a protein called PSIP1. This protein is an important RNA-binding protein and transcriptional coactivator, and its related pathways include activation of nervous system and cAMP-dependent PKA. We reviewed the previous literature and found that PSIP1 protein is closely related to tumor stemness and drug resistance, but PSIP1 has not been studied in HCC. Germline polymorphisms in PSIP1 enhancers are closely associated with progression-free survival in epithelial ovarian cancer, and also with carboplatin and paclitaxel response to drug therapy[66]. The LEDGF/P75 integrase-binding domain interactome contributes to the survival, clone formation, and tumor sphere formation of docetaxel-resistant prostate cancer cells[67]. The overexpression of LEDGF/P75 is associated with tumor invasive characteristics, including resistance to cell death and chemotherapy, and targeted stress of LEDGF/p75 can sensitize chemotherapy-resistant prostate cancer cells to taxanes[68]. Another study has shown that PSIP1/P75 plays an important role in the plasticity of neural crest-derived melanoma and is an important marker of stemness of melanoma[69]. Based on the relevant information of PSIP1 in the GEPIA database and our cell function experiments, we found that the high expression of PSIP1 in HCC promoted the progression of HCC and lenvatinib resistance, and was significantly correlated with the disease-free survival and overall survival of HCC patients. Then, through the rescue experiment, we found that interfering with the expression of PSIP1 could partially reverse the promotion and improvement of circRNA-mTOR on the proliferation ability, clone formation ability, invasion and metastasis ability, tumor stemness, and lenvatinib IC50 value of HCC, and it can partially reverse the inhibitory effect of circRNA-mTOR on cell apoptosis.
In the study of Cai et al., circRNA-SORE in sorafenib resistant HCC cells could bind to the main oncogenic protein YBX1 in the cytoplasm, thereby preventing the interaction between YBX1 and E3 ubiquitin ligase PRP19, and then blocking the PRP19-mediated degradation of YBX1, thereby mediating the enhancement of sorafenib resistance in HCC[8]. cIARS (hsa_circ_0008367) is a highly expressed circRNA in HCC cells after sorafenib treatment, which can mediate autophagy inhibition through interaction with RNA-binding protein ALKBH5, and positively regulate sorafenib induced iron death[70]. These studies suggest that circRNAs can change the total amount of RBP protein by stabilizing protein in the context of drug resistance of HCC. Therefore, we first explored the effect of overexpression or interference of circRNA-mTOR expression on the total amount of PSIP1 protein, but the experimental results showed that the total amount of PSIP1 protein did not change significantly after overexpression or interference of circRNA-mTOR expression. In other words, circRNA-mTOR did not act by increasing or decreasing the expression of PSIP1. So, we went on to investigate the literature and found that circRNA can act by altering the intracellular localization of RBPs in HCC. One study found that M6A-modified hsa_circ_0058493 promotes its intracellular localization from nucleus to cytoplasm by binding to RNA binding protein (YTH Domain-Containing Protein 1), thereby accelerating the progression of HCC[71]. In addition, one study has shown that CircBACH1 (hsa_circ_0061395) promotes the translocation of HuR from the nucleus to the cytoplasm by binding to RNA-binding protein (HuR), thereby eliminating the inhibitory effect of circBACH1 on p27 translation and promoting the growth of HCC[72]. In view of this, we experimentally detected the localization of circRNA-mTOR and PSIP1 in the nucleus and cytoplasm, and the results showed that circRNA-mTOR could promote the translocation of PSIP1 from cytoplasm to nucleus. To further explore the downstream pathway of hsa_circ_0009792/PSIP1, we first analyzed the proteins interacting with PSIP1 in the STRING database (https://cn.string-db.org). The results showed that MYC (c-Myc) could bind to PSIP1 and had the closest relation with PSIP1. A query of Gene Cards database revealed that c-Myc is a proto-oncogene encoding a nuclear phosphoprotein that plays a role in cell cycle progression, apoptosis and cell transformation, and high expression of this protein is often observed in many human tumors, and it is a regulator of somatic reprogramming which plays an important role in controlling the self-renewal of embryonic stem cells. A study has shown that circRNA UBAP2 could increase HCC tumorigenesis by upregulating c-Myc expression[73]. Hsa_circ_0091581 promoted the proliferation of HCC cells by blocking miR-526b from degrading c-MYC mRNA[74]. Another study found that circVAMP3 interacts with CAPRIN1 and G3BP1 to trigger the phase separation of CAPRIN1 and promote the formation of stress particles in cells by acting as a molecular scaffold, and through stress particles, circVAMP3 inhibits c-Myc translation and thus exerts its tumor suppressor properties[75]. In the study of Shinohara et al., PSIP1 can promote the expansion of tumor stem cells and induce drug resistance by activating c-Myc oncoprotein[76]. In another study, PSIP1 protein was found to co-immunoprecipitate with the transcription factor c-Myc in docetaxel-resistant prostate cancer cells, and depletion of LEDGF/p75 and selected interacting partners robustly decreased the survival, clonogenicity, and tumor sphere formation capacity of docetaxel-resistant cells[67]. These studies suggest that circRNAs and PSIP1 can affect tumor stemness or drug resistance by affecting c-Myc. We also found that the expression of c-Myc and tumor cell stemness markers (Oct4, Nanog and SOX2) increased after overexpression of circRNA-mTOR. This is also consistent with our previous finding that circRNA-mTOR can play a biological role by increasing the stemness level of HCC.
Recent study has demonstrated that the inhibition of Fibroblast Growth Factor Receptor (FGFR) by lenvatinib treatment leads to feedback activation of the EGFR-PAK2-ERK5 signaling axis, which is blocked by EGFR inhibition[6]. In other words, EGFR activation is the main cause of lenvatinib resistance in HCC. Moreover, a number of previous studies have confirmed that there is a large number of associations between the mTOR pathway and the FGFR pathway: the homeostasis of intestinal stem cells requires CDC42 to maintain through the activation of downstream YAP-EGF-mTOR[77]; the overactivation of mTORC1 in cancer can mediate the downstream RUNX1/EGFR pathway to affect STAT activation, and the expression of mTOR in a variety of human cancers is positively correlated with EGFR, which fully indicates the interaction between them[78]; the activation of EGFR/mTOR pathway directly affects apoptosis and iron death in ovarian cancer[79]; a glioma related article showed that Rictor, a key molecule in mTORC2 complex, could directly up-regulate EGFR protein expression[80]. Based on these related studies, we also examined the effect of circRNA-mTOR on the changes of EGFR in HCC cells, and the results showed that circRNA-mTOR could up-regulate the expression of EGFR. This suggests that the regulation of EGFR expression by circRNA-mTOR via related protein bypass may also be one of the mechanisms mediating lenvatinib resistance in HCC, but this still needs further experimental verification.
In conclusion, our study suggests that circRNA-mTOR plays an important role in the maintenance of lenvatinib resistance in HCC (Fig. 7F). CircRNA-mTOR can affect the nuclear translocation of PSIP1 by binding to it, and promote the progression and lenvatinib resistance through the PSIP1/c-Myc axis in HCC. In addition, the regulation of EGFR expression by circRNA-mTOR via related protein bypass may also be one of the mechanisms mediating lenvatinib resistance in HCC. Together, this study provides a certain experimental basis for the targeted drug therapy of HCC, and puts forward new ideas, new insights and new methods in understanding the occurrence and development of HCC, and it is of great significance to seek new markers and targets for the diagnosis and treatment of HCC and reduce drug resistance. Of course, this study still needs more clinical samples, especially drug-resistant HCC cells and tissue samples before and after lenvatinib treatment. At the same time, the mechanism of EGFR bypass regulation still needs to be further explored.
{kind=link}