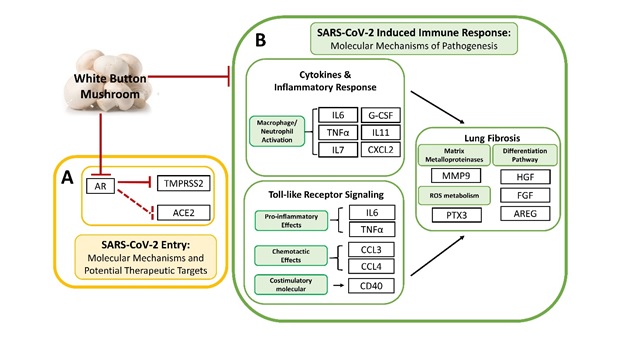
TMPRSS2 and ACE2 are two putative receptor proteins for the virus, SARS-CoV-2, to gain entry into host cells [9]. TMPRSS2 was first characterized as an androgen-regulated gene in the prostate gland [10, 11], but emerging studies have recently demonstrated that TMPRSS2 is also positively regulated by androgen in murine lungs and in human lung cells [12]. Since TMPRSS2 is implicated in COVID-19 pathogenesis, it supports a promoting role of androgens that males, relative to females, are disproportionately affected by COVID-19 in terms of mortality and morbidity [8]. Therefore, repurposing anti-androgenic drugs in the context of the COVID-19 pandemic is one of the major efforts currently being made [13]. Several preclinical studies, including some employing large epidemiological cohorts, suggested that blocking androgen signaling might protect against COVID-19 [33, 34]. The clinical trial using AR antagonists, proxalutamide/GT0918, in COVID-19 is currently underway (ClinicalTrials.gov, NCT04446429. Anti-Androgen Treatment for COVID-19). The trial released the preliminary analysis of proxalutamide as a treatment for COVID-19 patients. The data showed that proxalutamide could significantly ameliorate symptoms and prevent hospitalization for COVID-19 patients [35]. Putative use of WBM in mitigating COVID-19 was supported by findings from our clinical phase 1 trial in prostate cancer patients, as well as from the very recent preclinical studies that address the anti-androgen receptor activity of WBM. Our clinical phase I trial in prostate cancer patients indicated that oral dietary WBM suppressed circulating PSA levels [6]. We recently showed that dietary WBM antagonized DHT-induced AR activation and PSA expression in prostate cancer models and mouse prostate glands [5]. Considering these findings from our clinical and preclinical studies, dietary WBM could antagonize DHT-induced AR activation and TMPRSS2 expression throughout the body, including the putative COVID-19 targeted organs of lungs, small intestine, and kidneys.
In the current study, we demonstrated that androgen regulates the expression of AR and TMPRSS2 in subsets of pulmonary and intestinal epithelial cells. The AR and TMPRSS2 levels are markedly elevated in the lungs and the intestine upon DHT exposure, while WBM and an AR antagonist, enzalutamide, effectively repressed DHT-induced transcriptions of AR and TMPRSS2 (Figs. 2 and 3). To date, there are debates regarding the androgen-induced TMPRSS2 regulation in lung epithelial cells. A preprint article suggested no evidence for increased TMPRSS2 expression in the lungs of males, compared to females, in humans and mice. In their male mouse model, treatment with enzalutamide did not decrease pulmonary TMPRSS2 [36]. However, several other studies demonstrated with strong evidence that androgen regulates the expression of TMPRSS2 and AR in subsets of lung epithelial cells, and AR antagonists inhibit SARS-CoV-2 infections in vitro [37, 38]. Another preprint article also demonstrated the co-expression of AR and TMPRSS2 in specific lung cell types. Treatment with enzalutamide reduced TMPRSS2 levels in human lung cells [39]. As compared to the numerous investigations of AR-TMRPSS2 in lungs, very few research studies document this regulation in the other putative COVID-19 target organs, such as the small intestine and kidneys [40]. There was one preprint article that noted the reduced TMPRSS2 staining in the bronchial epithelium of the lungs, columnar epithelium of small intestine, and proximal convoluted tubules in the kidneys of androgen-deprived C57BL/6 mice from castration [41]. In our experiments, we observed that AR and TMPRSS2 levels were significantly elevated in the lungs and the small intestine upon DHT exposure, while intake of WBM and enzalutamide effectively repressed DHT-induced transcriptions of AR and TMPRSS2 in multiple organs. Our study using a male mouse model provided additional evidence that AR induced TMPRSS2 expression in the putative COVID-19 targeted organs of lungs, small intestine, and kidneys. By the same principle as the ongoing clinical trial with AR antagonists for COVID-19, our studies suggest WBM to be a potential dietary-based intervention via AR-TMPRSS2 transcriptional inhibition of critical host factors in the treatment or prevention of COVID-19 at virus entry level.
In terms of immune pathogenesis, the severity and clinical outcomes of COVID-19 patients are due to not only the viral load, but also to the host’s response that is triggered by viral entry and replication [17]. COVID-19 infection is characterized as a pro-inflammatory status by high levels of inflammatory factors produced by hyperactive immune cells. The inflammatory factors include cytokines such as IL-6, TNF-α, and G-CSF and chemokines such as CCL2, CCL3/4, and CXCL-2. These, together with reactive oxygen species, have been recognized to induce acute respiratory disease syndrome (ARDS), leading to lung fibrosis and possibly death [15, 16]. Therefore, systematically alleviating this hyper-activated inflammatory state is crucial to improve the prognosis and outcome of COVID-19 [17]. Several approaches such as IL-6 inhibitors and immuno-checkpoint inhibitors have been proposed to counteract the cytokine storm present in COVID-19 patients. However, the benefits, dose, and duration of these approaches remains to be validated [42]. A massive infiltration of mononuclear cells has also been detected in infected lungs, with parallel low levels of hyperactive T cells in circulation [16]. The immense migration of innate immune cells to the infected tissue, in order to control the viral replication, could contribute to the tissue damage and lead to multiple organ failures [43]. The immune system develops multiple mechanisms in order to control the excessive immune activation, including induction of an inhibitory receptor, production of anti-inflammatory factors, and expansion of regulatory cells, et al [44]. Meanwhile, MDSCs are a group of regulatory cells known to have the remarkable capability to regulate inflammatory responses and suppress T cell responses [45]. MDSCs have also been described to be highly increased in COVID-19 patients [31]. An emerging study further revealed that PMN-MDSCs expanded during the early stages of COVID-19 and were correlated with IL-1β, IL-6, IL-8, and TNF-α plasma levels, particularly in patients who required intensive care treatments, suggesting new therapeutic options geared towards MDSCs [32].
Emerging discussions and studies are referring to β-glucan as an efficient, low-cost, and safe way to overcome the hyper-inflammatory status while balancing effective immune responses [46, 47]. In fact, β-glucan has been widely shown to exert antiviral properties and decrease the severity of both upper and lower respiratory tract viral infections in both animal and human studies [48, 49]. Following exposure to β-glucan, innate immune cells undergo reprograming that results in immune enhancement by the activation of NK cells and CD4+ Th1 cells and suppression of the inflammatory response via downregulation of pro-inflammatory factors such as IL-6, CCL2, CXCL10, et al. [23]. β-glucan was reported to enhance anti-cancer immunity by suppressing MDSCs [24, 25]. Our clinical phase I trial in prostate cancer patients also indicated that oral dietary WBM reduced the counts of MDSCs [6]. In the line of evidence, we hypothesized that dietary WBM may display integrative immunoregulatory effects by suppressing pro-inflammatory cytokines, as well as MDSCs, through β-glucans.
To test our hypothesis, we demonstrated (Fig. 4) that WBM or LT predominantly suppressed a panel of cytokines when compared to Control. The cytokine signatures associated with treatments of WBM and LT greatly overlapped (15 out of 26 for WBM, 15 out of 15 for LT). More importantly, the unbiased GSEA analysis suggested that the shared cytokines included molecules that were associated with the Lung Fibrosis Pathway and the COVID-19 Adverse Outcome Pathway. These cytokines, such as IL-6, IL-7, CCL-2/MCP-1, CCL-3/CCL-4 MIP-1α/β, TNF-α, and G-CSF, have been implicated in COVID-19 pathogenesis [29, 30]. As expected, when we characterized MDSCs in both blood and spleen of WBM or LT-treated mice (Fig. 5), we observed the regulatory trend that both WBM and LT decreased the total counts of M-MDSCs and PMN-MDSCs, in peripheral blood and spleen. Such evidence supports the fact that β-glucan is the major component in WBM, acting as an immunomodulator. Besides, we also observed a subset of 11 factors which was selectively regulated by WBM. Among these, we pointed out a defined cytokine trait (CCL-12/MCP-5, CCL-19/MIP-3β, CXCL-2/MIP-2, et al) of severe COVID-19 outcome [30]. In considering that WBM is a mixture of multiple components, we cannot rule out the possibility that additional chemicals beside β-glucan in WBM may potentially exert immunoregulatory activities.
In conclusion, COVID-19 is a respiratory and systemic disorder accompanied by SARS-CoV-2 entry into host cells and rapid replication. Virus infection triggers immune dysregulation and a cytokine storm, ultimately leading to a range of symptoms along the clinical spectrum that include asymptomatic or mild respiratory symptoms, severe lung injury, multi-organ failure, and death [14 ~ 17]. Although specific target molecules and agents can act on each step of pathogenesis, in considering the complexity of the immune system and its systemic response, such interventions may be efficacious but come along with adverse reactions [50]. WBM is composed of a variety of chemical ingredients. Our preclinical studies have revealed that WBM is a unique food that can suppress TMPRSS2 expression through its anti-androgenic activity mediated through conjugated linolic acid [5] and promote an anti-inflammatory response, possibly by β-glucan. We hereby propose WBM consumption as a potential efficient, low-cost, and safe dietary approach to mitigate COVID-19. Our completed phase 1 trial using WBM determined that a dose level of up to 14 g WBM powder (equal to 140g fresh WBM)/d resulted in minimal side effects, mostly limited to grade 1 abdominal bloating. Therefore, WBM intervention is considered safe with a demonstrated mean compliance of 98.6% [6]. However, clinical studies and trials will be needed to prove its efficacy against COVID-19.
{kind=link}