In experimental studies, Morin has shown protection against cardiovascular, cerebrovascular, diabetes, liver, and kidney diseases through antioxidant, antiapoptotic, and anti-inflammatory mechanisms. However, the effect of morin on myocardial injury in diabetic conditions is still unknown. In the present study, the effect of Morin on ISO-induced myocardial ischemia in diabetic rats was evaluated. In the results, we found that Morin reduced normalized blood glucose and serum insulin levels, strengthened the antioxidant defense system, preserved myocardial architecture, and prevented the increase in inflammation and apoptosis in the myocardium. This could be due to the regulation of multiple signaling pathways, i.e. AGE-RAGE/Nrf2/HO-1, MPAK, and GSK-3β/Akt/eNOS.
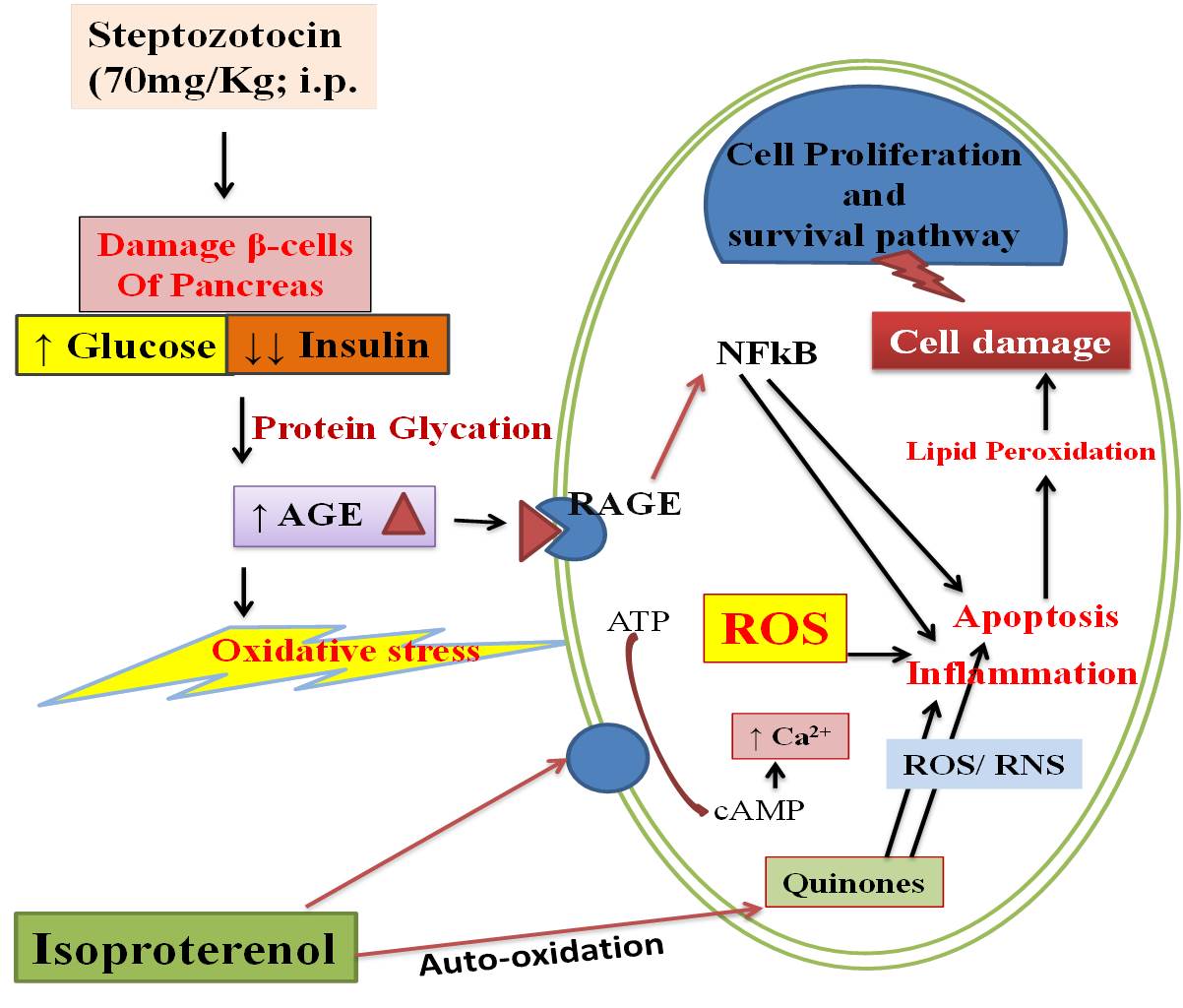
Streptozotocin is an antibiotic that is widely used to produce diabetes in experimental animals. It enters the pancreatic β cells via GLUT2 transporter. In β cells, it has a cytotoxic effect, causing DNA fragmentation, depletion of intracellular NAD+ and ATP, and mitochondrial dysfunction, which ultimately leads to necrosis of pancreatic β cells. A decrease in the number of pancreatic β cells produces a relative or absolute decrease in insulin levels resulting an increase in blood glucose levels (Szkudelski 2012). A decrease in insulin level leads to the breakdown of proteins, and fatty acids, and increased muscle wasting which reflects a decrease in body weight (Cheng et al. 2013). Similarly, in this study, we found that an increase in hypo-insulinemia, hyperglycemia, and a decrease in body weight in diabetic group animals and Morin treatment significantly reversed (but not completely) their levels.
It has been known that hyperglycemia activates the production of AGE, which then interact with RAGE and enhance the production of free radicals (Daffu et al. 2013).When diabetic rats were further subjected to isoproterenol, it leads to more production of free radicals and depletion of the antioxidant defense system. There, free radicals further react with lipids, proteins, and DNA and cause cell membrane damage. This results in lipid peroxidation and the release of cardiac injury markers (CK-MB & LDH) from the cell membrane. Previously, various studies have shown that natural antioxidants protect against myocardial injury in diabetic rats by inhibiting the AGE/RAGE pathway (Suchal et al. 2017; Thakur et al. 2021). With increased oxidative stress, the body activates the antioxidant system which prevents the generation of free radicals, lipid peroxidation, and depletion of antioxidants. During investigations, we found that towards protection from free radicals, the Nrf2 pathway was activated as shown by other researchers. On activation, Nrf2 binds to the antioxidant response elements (ARE) and increases the production of various antioxidant enzymes (David et al. 2017; Zhang et al. 2019). In line with this, we extended the analysis and found increased levels of AGE/RAGE proteins, MDA (a marker of lipid peroxidation), cardiac injury markers, and decreased antioxidants (GSH and SOD) in the DC and D+I groups rats. On the other hand, treatment with Morin reduced the AGE/RAGE levels, lipid peroxidation, LDH, and CK-MB levels and increased the antioxidant status, which could be due to the activation of Nrf2 pathway. Similar to this, previous studies have demonstrated the antioxidant role of Morin in various disease models (Ozdemir et al. 2020; Khamchai et al. 2020; Verma et al. 2020; Sang et al. 2017).
To check the effect of Morin on apoptosis in diabetic myocardium, various antiapoptotic and proapoptotic protein levels were measured. The intrinsic pathway of apoptosis involves the release of cytochrome c from mitochondria to the cytosol where it binds to Apaf-1 and causes activation of the caspase-dependent apoptotic pathway. Bcl-2 can inhibit apoptosis by preventing the release of cytochrome c from the mitochondrial membrane. Puma and Noxa are two members of the Bcl-2 family which are involved in apoptosis and have been found to be stimulated by p53. Both intrinsic and extrinsic pathways of apoptosis end at the executioner phase of apoptosis. Caspase- 3, 7, and 9 are members of the executioner phase of apoptosis which further cleaves various cytokines such as PARP leading to biochemical and morphological changes in the apoptotic cells (Elmore 2007). Previously, it has been found that Hsp 70 and 27 act at different steps of the intrinsic pathway of apoptosis and prevent apoptosis (Paul et al. 2002; Bratton and Salvesen 2010). In our study, there was a decreased level of antiapoptotic proteins (Hsp 70, 27, and Bcl-2) and increased levels of proapoptotic markers (cytochrome c, Bax, caspase-3, PARP, and p53) in DC and D+I group rats and was reverted in the Morin treatment group. In agreement with the findings, various other studies have shown the antiapoptotic potential of Morin in different tissues/ organs (Khamchai et al. 2020; Chen et al. 2017; Liu et al. 2018). Furthermore, it has been documented that Akt/GSK-3β signaling pathway regulates apoptosis and promotes cell survival, while phosphorylation of Akt has been shown to reduce apoptosis (Li et al. 2016). In line with this, we found increased levels of Akt in the Morin treatment, which might be responsible for the decreased levels of proapoptotic proteins in the treatment group. Rizvi et al. have shown the protective effect of Morin through the activation of Akt/GSK-3β pathway (Rizvi et al. 2015).
It has been well-documented that sustained oxidative stress can lead to the activation of the MAPK pathway. MAPK pathways consist of three proteins, ERK, JNK, and P38. Activation of JNK and P38 has been shown to induce inflammation in the myocardium by stimulation of transcription factors such as NF-κB/IKKβ (Wu et al. 2019). In the inactive form, NF-κB resides in the cytoplasm with the inhibitory protein IκB. In response to stressed conditions, there is phosphorylation of IκB by IKK which causes nuclear translocation of inflammatory cytokines, i.e., TNF and IL-6. HMGB1 has also been shown to increase the production of inflammatory cytokines by interacting with RAGE and toll-like receptors (Zhang et al. 2018). In this study, there was an increased level of MAPK, NF-κB/IKKβ, HMGBI, and inflammatory cytokines in the DC and D+I group rats, which were significantly reduced by Morin treatment. Recent studies have shown that Morin can act as an anti-inflammatory agent by inhibition of inflammatory pathways (Khamchai et al. 2020; Chen et al. 2017).
Recently, the role of inflammasomes has been extensively studied in myocardial ischemic injury, Alzheimer’s disease, and diabetes (Guo et al.2015). NLRP3 inflammasome consists of NLRP3, cysteine protease pro-caspase-1, and the adaptor protein apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC). In response to oxidative stress, NLRP3 forms a complex with ASC which results in caspase-1 activation. Activated caspase-1 induces the cleavage of IL-1β and IL-18 to their active form, which causes inflammation in the tissue. Activation of caspase-1 also leads to inflammation-related cell death known as pyroptosis (Takahashi 2019). In the present study, there was an increased level of inflammasomes (NLRP3, IL-1β, and caspase-1) in the DC and D+I groups and treatment with Morin reduced levels of inflammasome proteins. Li et al. have shown that morin protects against Listeria monocytogenes induced infection by inhibition of the inflammasome pathway (Li et al. 2020).
{kind=link}