Diatoms mostly rely on the movement of the water to move them; they are immobile in most situations. However, some diatoms have a slit in their frustule called a raphe that allows them to glide down a surface. Pennate Raphid diatoms are notable for their motility and exhibit distinctive movements that are directly tied to the structure of their raphe system. (Round et el., 1990). The raphe is covered by actin filament. The rapid pennate's ability to move depends on the actin filaments that support it. It is obvious that blocking actin co-ordinately affects movement, even though the exact role played by actin filaments in motility is still not entirely known. Actin is undoubtedly important, even if it may be utilized for mucilage insertion, direction, or connection to motor protein force-generating. In motile raphid diatoms, the actin cables, which are made up of big bundles of actin filaments, appear to be crucial for active, properly controlled motility. However, nothing is known about the relationship between the cables' filaments and the fibers of raphe mucilage. This enables them to exude mucus that adheres to a surface, but how does that mucus aid in their mobility? We are unsure. Although we can observe that they can move and that mucus is always left in their wake when they do so, the precise method by which they do so is unknown to us.
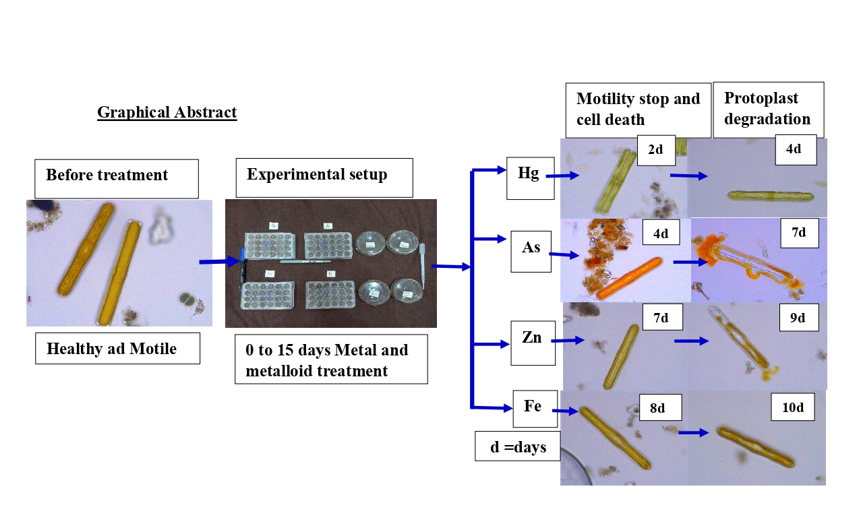
Our study, which is derived from experiments performed on six diatom genera, has shown that cells treated with metals (Hg, As, Zn, and Fe). We measured the proportion of cells moving Vs. not moving to determine the effect these metals’ concentration has on diatom motility. Iron and zinc are essential metals for life, whereas Arsenic and Mercury are not essential metals for life (Allan., 1997). All six diatomic genera showed a tremendous increase in % motile cells at the high concentration of zinc and iron, thereby indicating remarkable tolerance to zinc and iron. In contrast, all six genera were more abundant motile cells at low Arsenic concentrations than at the high arsenic concentration. (Bonilla et al.2005) says through his study on metal pollution. N. Palea is a metal-tolerant species that was found in abundance in the pollution from the mine spill and was infected with the mine spill metal pollution.
Mercury (Hg)
Mercury, a metal produced due to the increase in anthropogenic emissions, has become a serious pollutant to water and land ecosystems and human health (Nagajyoti et al. 2010). Mercury is soluble, highly reactive, and can accumulate in higher plants and aquatic organisms (Nagajyoti et al. 2010). It is similar to zinc (which is a nutrient of algae) and instead participates in metabolic activities (Kuper & Andresen 2016). It also harms algae. Potential role of actin and myosin in diatom cell motility, (Poulson et al., 1999) found in their experiment that actin, which is a cytoskeleton system, plays an important role in motility. His experiment took three different concentrations of latrunculin, an anti-actin drug, and observed that these concentrations were the least necessary to stop cell motility within 30 minutes. Butanedione monoxide (an antimyosin drug) also lost diatom motility, suggesting that actin–myosin proteins are essential for diatom motility. (Khan et al., 2017) found in their study that exposure to MeHg, a form of mercury, can epigenetically modify MMP9 to promote cytoskeleton disruption leading to loss of kidney function, Oxidative stress of metal pollution produces free radicals or reactive oxygen species (ROS), the effects of which on cellular redox status (Pinto et al. 2003). We have not found any previous studies on how mercury affects the cytoskeleton system of algae, but as we observed in our study, the motility was left at 2–5% within 24 hours. This suggests that mercury also has a profound effect on the cytoskeleton system of algae. Further studies are needed to look at the effect of mercury on the cytoskeleton system of algae in the future.
Arsenic (As)
In the natural ecosystem, arsenic is a widely dispersed metalloid. Arsenic is a chemically similar element to phosphorus because arsenic is found just below phosphorus in the periodic table and both have the electronegativity almost same and the same atomic radius. Phosphate, a crucial mineral for algae, is an analog of arsenic, Algae can accumulate arsenic inside themselves. which can reduce the growth of biofilm in streams (Barral-Fraga et al. 2015). (Barral-Fraga et al. 2015) explains the toxic effect of arsenic on biofilm communities' reduced diatom cell size. The toxic effect of arsenic comes because it interferes with the metabolism of phosphorus. Phosphate participates in a variety of biological activities like ATP, ADP, phospholipids, and ion pumps may be a subsequent synthesis of phosphate transport and depend on phosphate availability in the environment (Miot et al.2009). We know that the cytoskeleton system plays an important role in cellular motility. Actin is a cytoskeleton filament ATP is required for the polymerization of actin. If phosphorous is replaced by arsenic, actin will not be able to polymerize, which will have a profound effect on cell motility. ATP (Adenosine Tri Phosphate) also called energy coin, which gives energy for the existence of any cell. If the cell does not receive energy and is unable to perform its metabolic and physiological activities properly, the cell may even die. We also see the motility inhibitory effect of As on diatomic genera (Navicula, Nitzschia, Amphora, Achnanthes, and Pinnularia). We found 0% motility within 4 days after high-concentration arsenic treatment in Navicula and 10% motility in medium and low arsenic concentrations. In the case of Nitzschia lose their motility completely in 6 days of treatment with arsenic high concentration and 10% motility in medium and low As concentrations. While the motility in control was observed for 10–12 days, that means arsenic may give a toxic effect on the microalgae. In contrast to diatoms (Simon et al. 2010) work on a bacterium called Halomonadaceae strain GFAJ-1 was found in Mono Lake, California, and can develop by replacing phosphorus with arsenic. a Gamma proteobacterium belonging to the Halomonadaceae family has grown aerobically with AsO4 3–, glucose, and no additional PO4 3– (+ As/–P condition). Members of this family have previously been demonstrated to collect As within their cells. This probably happens when the amount of phosphorus is either low in the environment or phosphorus is found less than the amount of arsenic. Arsenic is toxic to freshwater microalgae at high concentrations, particularly at a low ambient concentration of phosphate.
Zinc (Zn)
Zinc is an essential nutrient for growth that acts as a cofactor for more than 300 enzymes and is involved in almost all metabolic reactions (Anderson et al., 1978; Vallee and Auld, 1990). Zn also serves as cofactors for enzymes participating in CO2 fixation enzyme carbonic anhydrase (Moroney et al. 2001), in RNA polymerase, and also in alkaline phosphatase (Sunda et al.,2012). Although zinc is an essential metal for the growth of diatoms and other microalgae, it can prove dangerous to aquatic organisms if it is found in excess or less than its permissible limit. Diatoms use dissolved Si to build their cell wall through biomineralization. Several studies also demonstrated decreased Si uptake under Zn deficiency (Rueter and Morel, 1981; De La Rocha et al., 2000; Franck et al., 2003). We have seen in our experiments that Achnanthes, Nitzschia, and Pinnularia show tolerance to zinc and iron in 40–50% of live cells for about 14 days. Whereas Navicula, Amphora, and Diplonais were found to be very sensitive to zinc, 14-day incubation of zinc enrichment enhanced the relative abundance of species like Achnanthes exigua and Nitzschia amphibia and these were metal (Zn) tolerant (Pandey and Gaur. 2014). Green algae and diatoms dominated the local algal population. In comparison, copper effluents in the khetri region, and zinc smelter effluents had a higher concentration of green algae. Green algae and diatoms dominated the zinc community. The relative abundance of Achnanthes exigua, Nitzschoid diatoms, and green algae was decreased at the sites with severe pollution when compared to less polluted areas, where four diatom species (Achnanthidium minutissimum, F. capucina, Pinnularia conica, and U. ulna) predominated (Pandey and Bergey 2016). We also found the same effect of Zn in our experiment. Zn work as a nutrient for the diatoms. We found a high % motile cells under medium Zn concentration in comparison to low and high Zn concentrations. Although zinc is essential in the normal functioning of living diatoms concentration beyond the permissible limits causes death for diatoms because of their intracellular accumulation nature (Pandey and Gaur. 2014).
Iron (Fe)
Fe is necessary for the development of the nitrate reductase enzyme complex as well as for photosynthesis, carbon fixation, and other processes. Diatoms lacking in Fe have significantly lower rates of carbon fixing and nitrate absorption. (Geider and La Roche,1994). Iron is the most significant trace metal in terms of quantity since it is a component of the Photosystem II complex (PSII), which requires two iron atoms per subunit, the Cytochrome b6f complex, which requires five iron atoms per monomer, the Cytochrome c6 complex, which requires one iron atom per monomer, the Photosystem I complex (PSI), which requires twelve iron atoms, and ferredoxin (two iron atoms), (Raven et al.1999., Strzepek et al.,2012., Xia Gao, Chris Bowler, and Elena Kazamia.2021). The community structure of species in the HNLC (high nutrient, low chlorophyll) zone is also influenced by the iron shortage (low concentration), which also hinders the establishment of large species with high half-saturation constants for Fe uptake (Gall et al., 2001; Sedwick et al., 2000; Timmermans et al.,2001). Just as there are many disadvantages due to iron deficiency, similarly a major drawback of iron excess is Fe-induced diatom blooms, which were seen during two in situ fertilization experiments carried out in the Western Sub-Arctic Pacific (SEEDS, 2001) and Eastern Sub-Arctic Pacific (SERIES, 2002). These experiments confirmed that the HNLC waters were indeed Fe-limited. (Tsuda et al., 2003; Crawford et al., 2003; Boyd et al., 2004). iron acts as a nutrient for microalgae. In our experiment, we found that the treatment of iron kept the motility in the diatom for a long time. In contrast, Hg and As. In the diatom Phaeodactylum tricornutum, exposure to Fe increased Chl content in a concentration-dependent manner (Kosakowska et al., 2004). We also observed in our experiment that diatoms were found to have more actives (motile) at higher concentrations of iron and were found to be active (motile) for a longer time.
{kind=link}