Preparation of GQDs and their uptake by T. brucei
GQDs were synthesized as illustrated in Fig. 1a. According to TEM images of the prepared GQDs (Fig. 1b), the core sizes of GQDs were 2.15 ± 0.59 nm (n = 100) with a relatively narrow size distribution. Despite their promising quantum yield and bleach-resistant autofluorescence, GQDs alone cannot be clearly imaged in the complex biological environment of T.brucei. Given these inadequacies, fluorescein fsothiocyanate (FITC), a typical fluorescent dye, was modified onto the surface of GQDs by EDC/NHS to enhance the fluorescence intensity of GQDs (Fig. 1a). The fluorescence excitation and emission spectra of GQDs or GQDs-FITC were shown in Fig. 1c. Ex/Em of 488/550 nm were chosen for GQDs-FITC to investigate the endocytosis of GQDs by T. brucei. The endocytosis system of T. brucei has been identified as a clathrin-mediated endocytosis (CME). CME is the most ancient known as mechanism of internalization in many lineages, and the involved protein participants have been known[40]. In T. brucei, CME is AP-2 independent and involves a cohort of lineage-specific clathrin-associated interactomes that drives CME. Clathrin is localized in the collecting tubules of flagellar pocket, where clathrin and its partners are recruited and assembled to perform endocytosis in T.brucei. The flagellar pocket is a particular type of plasma membrane invagination that surrounds the base of the flagella, and all trypanosoma endocytosis and exocytosis are polarized to this site as an only gateway to and from the host-parasite interface.
Although CME pathway focusing on small molecules as the nutrients has been investigated well in T. brucei, few reports are provided on nanoscale materials. After 6h of incubation, the obtained images visualized the phase contrast image of T.brucei cells, involving the blue nucleus stained with DAPI and the green GQDs-FITC (200 µg/mL) distributed in the cytoplasm of T. brucei cells (Fig. 1d). Pitstop2 is a classical inhibitor of clathrin-mediated endocytosis that are widely used in mammalian cells. This inhibitor has been shown to work well in T. brucei cells[24]. As shown in the last line of images, the pre-treatment of T. brucei cells with Pitstop2 caused a significant reduction of internalized GQD, indicating a key role of CME in GQDs uptake in T. brucei. The internalization of GQDs indicated the two-dimensional nanomaterials represented by GQDs can be used as promising nanomedicine materials for the diagnosis and treatment of parasitic protozoa.
Cytotoxicity induced by GQDs on T. brucei
Firstly, the cytotoxicity induced by GQDs in T. brucei cells was evaluated by MTS assay. As shown in Fig. 2a, the GQDs inhibited significantly the cell viability of T. brucei by a dose-dependent pattern. Half-maximal inhibitory concentrations (IC50) for GQDs after 24h incubation was 36.1 µg/mL, which was negligible toxic doses for mammalian cells[32]. In most cases, GQDS can cause significant toxicity in mammalian cells at doses between 150 and 300 µg/mL, suggesting GQDs can be applying as a potential anti-trypanosomic reagent or drug carrier in vivo. Next, the apoptosis/necrosis ratio of T. brucei cells treated with GQDs was assessed using the Annexin V-FITC/PI apoptosis assay. Flow cytometry (Fig. S1) and statistical analysis (Fig. 2b) demonstrated that GQDs mainly induced late apoptosis. In T. brucei, the apoptotic phenotypes have been concerned and described when the parasite was incubated with drugs[41]. However, unlike mammalian cells, there are no evidences that caspase-like activity on their respective substrates have been detected in pathogenic trypanosomatids. Up to now, the key factors contributing to this process including caspase genes, Bcl-2 family homologs and TNF-related receptor members have not been identified in T. brucei[42]. Instead, they have evolved other stress response mechanisms, such as prostaglandin D2(PGD2) treatment[43] and spliced leader RNA silencing (SLS) pathway.
Numerous reports have shown that apoptosis induced by both mitochondrial dysfunction and ER stress in trypanosoma are dependent on ROS, but independent of caspase-like proteases[44]. Oxidative stress is known to be the main pathway of apoptosis induced by nanomaterials, including GQDs. Thus, ROS production induced by GQDs was determined and results (Fig. 2c) showed that GQD treatment significantly increased the ROS level in T. brucei in a dose-dependent manner. The ultrastructural changes of subcellular organelles in T. brucei cell were significantly altered after exposure to GQDs, including a vesicular-like expanded endoplasmic reticulum (ER) and morphologically altered mitochondria (M) (Fig. 2d). In addition, a marked expansion of the flagellar pockets was observed. Usually, there is a significant increase in diameter and morphological defects following the occurrence of abnormal endocytosis or clathrin-associated protein depletion in flagellar pocket. In conclusion, T. brucei cells exposed to GQDs exhibited abnormal ultra-structural changes that were typical hallmarks of nano-induced toxicity.
Genome-wide target protein screening towards GQDs in T. brucei.
The ultra-small size, lamellar structure and oxygen-containing groups at the edges of GQDs enable them to interact specifically with protein molecules in T. brucei. Thus, identifying the key target proteins towards GQDs was a prerequisite for studying the cytotoxic mechanism induced by GQDs in T. brucei cells. A model of GQDs with a side length of 2.5 nm was constructed using Lerf-Klinowski model[45]. PDB files are provided in Supplementary Dataset S1. The 3D structures of 5190 proteins of T. brucei were obtained by three ways, including 283 proteins from PDB web, 3117 proteins using I-TASSER, and 1790 proteins using AlphaFold. The structures and thermodynamic parameters of the GQD-protein complexes were finally generated by molecular docking with the PatchDock program. PDB files of GQD-protein complexes were ranked according to binding strength which were provided in Supplementary Dataset S2. The top 200 proteins with high affinity to GQDs (Supplementary Dataset S3) were categorized into different groups based upon subcellular localization (Fig. S2), molecular function (Fig. S3), or biological process (Fig. S4). Results showed that a large proportion of GQD-binding proteins were located in mitochondria, endoplasmic reticulum, and membranes' integral components (Fig. 3b). The molecular functions of top 200 proteins could be mainly divided into two parts: transporter activity and oxidoreductase/ antioxidant activity (Fig. 3c).
Of the top 200 proteins (Supplementary Dataset S3), about one fifth had transporter activity (Fig. 3c), which involved important roles in a wide range of cellular activities such as nutrient uptake, metabolite release, and signal transduction. Representative target proteins with high affinity included: (1) glucose transporters[46] (Tb927.10.8440, Tb927.10.8490, Tb927.10.8530 and Tb927.10.8510) response to the maintenance of energy metabolism in T. brucei; (2) vacuolar ATPases (Tb927.10.200, Tb927.10.3760, Tb927.8.2100 and Tb927.11.7480) mainly distribute in the lysosome and acidocalcisomes of T. brucei, contributing to macromolecular degradation, nutrients storage and ion homeostasis[47]; (3) aquaglyceroporins (AQP1, Tb927.6.1520; AQP3, Tb927.10.14160) regulate osmosis and uptake of low molecular weight solutes such as glycerol and urea[48]; (4) mitochondrial carrier family (MCF, Tb927.10.14840, Tb927.9.4310) localize in the mitochondrial inner membrane, mediating the transport of various metabolic intermediates or maintain the mitochondrial ADP/ATP ratio in T. brucei[49, 50]. Thus, by interacting with transporters, GQDs affected the uptake, storage and degradation of nutrients, osmoregulation, ion homeostasis, energy metabolism and mitochondrial phosphorylation in T. brucei. As a result, GQDs could indirectly induce ROS production, and thus trigger trypanosoma apoptosis.
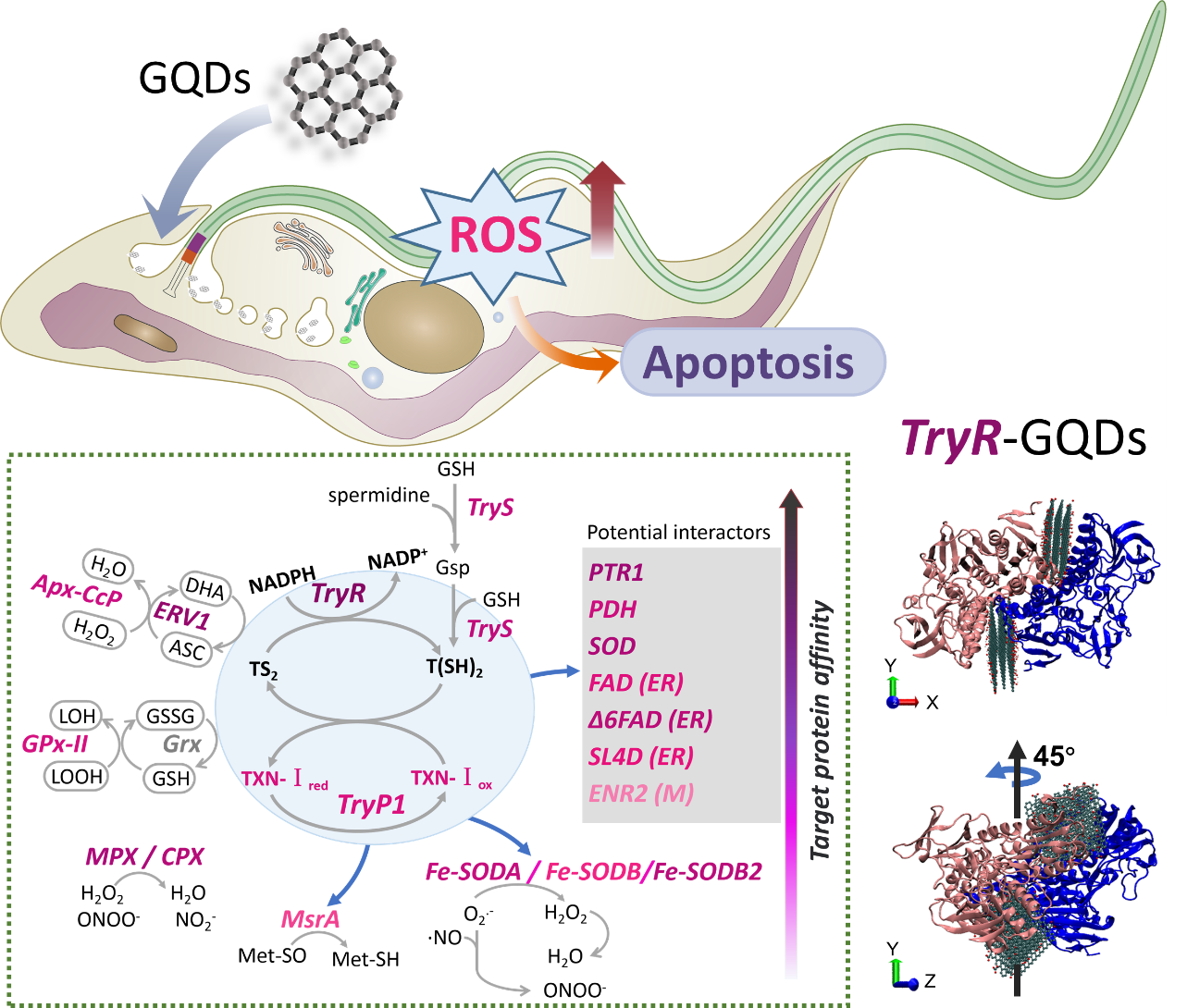
Strikingly, another large percentage of potential targets towards GQDs with high affinity were a group of enzymes associated with redox regulation activity (Fig. 3c). Trypanosomes are early-diverging eukaryotes that are susceptible to ROS, but lack the major disulfide reductases, such as glutathione reductase and thioredoxin reductase which control thiol-redox homeostasis in most living organisms. Trypanosoma have evolved a unique thiol-redox system based on trypanothione[51], which is condensation compound of GSH and spermidine synthesized by trypanothione synthetase (TS). The regeneration of reduced/oxidized trypanothione (T(SH)2/TS)2) are regulated by trypanothione reductase (TryR) using NADPH as co-factor[52]. During the thiol redox process, oxidized glutathione (GSSG), dehydroascorbate (DHA), and the species-specific dithiol protein tryparedoxin (TXN) can accept the reducing equivalents as intermediary molecules, and then (in their reduced state) transfer electrons to the antioxidant enzymes including peroxidases, reductases, glyoxalases, and transferases[53]. Docking simulation results (Supplementary Dataset S3) showed that up to 20 proteins related to the T(SH)2-TXN-NADPH system had high affinity with GQDS, suggesting that GQDs could directly disrupt the anti-redox system of T. brucei and thus initiate apoptosis.
Identification of TryR as vital target for GQDs in T. brucei.
Based on the T(SH)2-TXN-NADPH system, the species-specific antioxidant network consisting of enzymatic and nonenzymatic redox-active molecules, is responsible for the detoxification of ROS/RNS (H2O2, O2•−, •NO, ONOO−) in T. brucei. These redox-active enzymes located in distinct subcellular compartments in T. brucei have high susceptibility to oxidative stress, which are considered attractive candidates for drug targeting[54]. Results (Fig. S5) showed that GQDs could induce the ROS increase in a short time with a dose- and time-dependent relationship. Combined with the previous docking results (Fig. 3c), in which most antioxidant enzymes involved in T(SH)2-TXN-NADPH system had high affinity with GQDs, it was speculated that GQDs could interact directly with antioxidant enzyme molecules to trigger ROS-mediated apoptosis in T. brucei. To identify the crucial target proteins, MD simulation was performed using GROMACS software package to find out the details of interfacial interaction between GQDs and antioxidant enzymes (Fig. 4).
Twenty redox-active enzymes with high affinity towards GQDs were selected as potential targets to be investigated by MD simulations. Representative snapshots of nano-protein complex structures from MD simulations (Fig. 4a) provided the interfacial morphology between GQDs and enzymes. Bonding strength between GQDs and enzymes was measured using Gibbs free energy change (ΔG) obtained by averaging frames over 80–100 ns (Fig. 4b). Bonding strength consists of “ineffective bonding” and “effective bonding”. Ineffective bonding can be defined as a bonding that has little or no effect on enzyme activity. Effective bonding can be defined as a bonding that affects enzyme activity, and the bonding site is usually located at or near the enzyme activity center. Usually, an effective bonding event is the interaction with high bonding strength that occurs in the enzyme active center. In addition to bonding strength (Fig. 4b), the centroid distance between GQDs and the active site of the target protein was employed to identify effective bonding events (Fig. 4c). The active sites of enzymes were marked with the highlighted amino acids. Overall, TryR exhibited high interaction energy and the shortest centroid distance when binding to GQDs. TryR plays an extremely critical role in the trypanothione-dependent antioxidant network of T. brucei, by reducing TS2 to T(SH)2 with reducing equivalents provided by NADPH. In conclusion, TryR was identified as the most important target protein towards GQDs in T. brucei.
Validation of GQDs targeting TryR
To explore the inhibitory mechanism of the GQDs on TryR, the ultrastructures of the contact interface between GQDs and TryR were simulated by MD. According to the results (Fig. 5a), the strong interactions resulted from the hydrophobic interactions (VAL443, ILE339, PRO336, TRP21), hydrogen bonds (GLU18, ASN22, ASP116, SER109, THR335), and ionic bonds (ARG496, LYS108, LYS61) between GQDs and TryR amino acid residues. Among them, CYS52 and CYS57 residues are the components of TryR active site, meaning that GQDs can inhibit the activity of TryR by directly interacting with the amino acid residues of active sites in TryR. Considering that TryR functions as a dimer under physiological conditions, MD simulation method was further used to simulate the interaction between GQDs and TryR dimer. Results (Fig. 5b) showed that GQDs were inserted into the contact sites of TryR dimer to form a tight complex, where the activity center of TryR was completely blocked. The interaction energies between GQDs and TryR were calculated based on MD simulations (Fig. 5c), and the KD of 1.6×10− 8 M indicated a strong interaction between GQDs and TryR.
To validate the targeting effect of GQDs on TryR, biolayer interferometry (BLI) analysis was employed to quantify GQD-TryR interaction in vitro. His-tagged TryR recombinant (HIS-TryR) was expressed using a prokaryotic expression system and analyzed by SDS-PAGE (Fig. 5d). During BLI sensing, the HIS-TryR was first immobilized on the Ni-NTA biosensor, then immersed in the GQDs solution to capture the GQDs and subsequently dissociated in the dissociation solution (Fig. 5e). Dynamic characteristics of the GQD-TryR interaction based on BLI analysis were provided in Fig. 5f. The KD value (2.4×10− 8 M) obtained by BLI was comparable to that (1.6×10− 8 M) in MD (Fig. 5c), which also validated the accuracy of the previous theoretical simulation. To further verify whether GQDs affect TryR function, an enzyme activity evaluation system was developed by employing TS2 as catalytic substrate with the reducing equivalent provided by NADPH. The effect of GQDs on TryR enzymatic activity was evaluated by determining the absorbance decrease of NADPH at 340 nm. In the presence of TryR, NADPH (the maximum absorbance at 340 nm) will be converted to NADP+, which has no absorption at 340 nm. Considering the interference from GQDs absorbance themselves, a 3000-D ultrafiltration tube filter device was employed to remove the GQDs after interacting with TryR. Schematic diagram of the developed method and operation process was shown in Fig. 5g. The results (Fig. 5h) showed that GQDs exhibited a significant inhibitory effect on TryR activity. The inhibition rate reached 83.45% at 40 µg/mL of GQDs and 95.17% at 120 µg/mL of GQDs. In conclusion, GQDs could significantly inhibit TryR activity through strong interaction with TryR active site.
{kind=link}