Ameliorative effects of CNPs on experimental colitis
Encouraged by the nanozyme-like performance of CNPs, the therapeutic efficacy in the TNBS-induced colitis mouse model was investigated.
CNPs showed no obvious cytotoxicity incubation with HT-29 cells or RAW 264.7 cells for 24 h, indicating negligible cytotoxicity of CNPs at any tested concentrations (1.25-40 µM) (Fig. S4). Furthermore, hematoxylin and eosin (H&E) staining of the major organs of the administered mice (heart, liver, spleen, lung, kidney) revealed that CNPs exhibited no obvious toxicity at the administration dose (Fig. S5). We also conducted a hemolysis assay to evaluate CNPs hemocompatibility. The hemolysis rate demonstrated that CNPs had good hemocompatibility within 50 µM (Fig. S6). These results indicated that CNPs with good biocompatibility are promising for further biomedical applications.
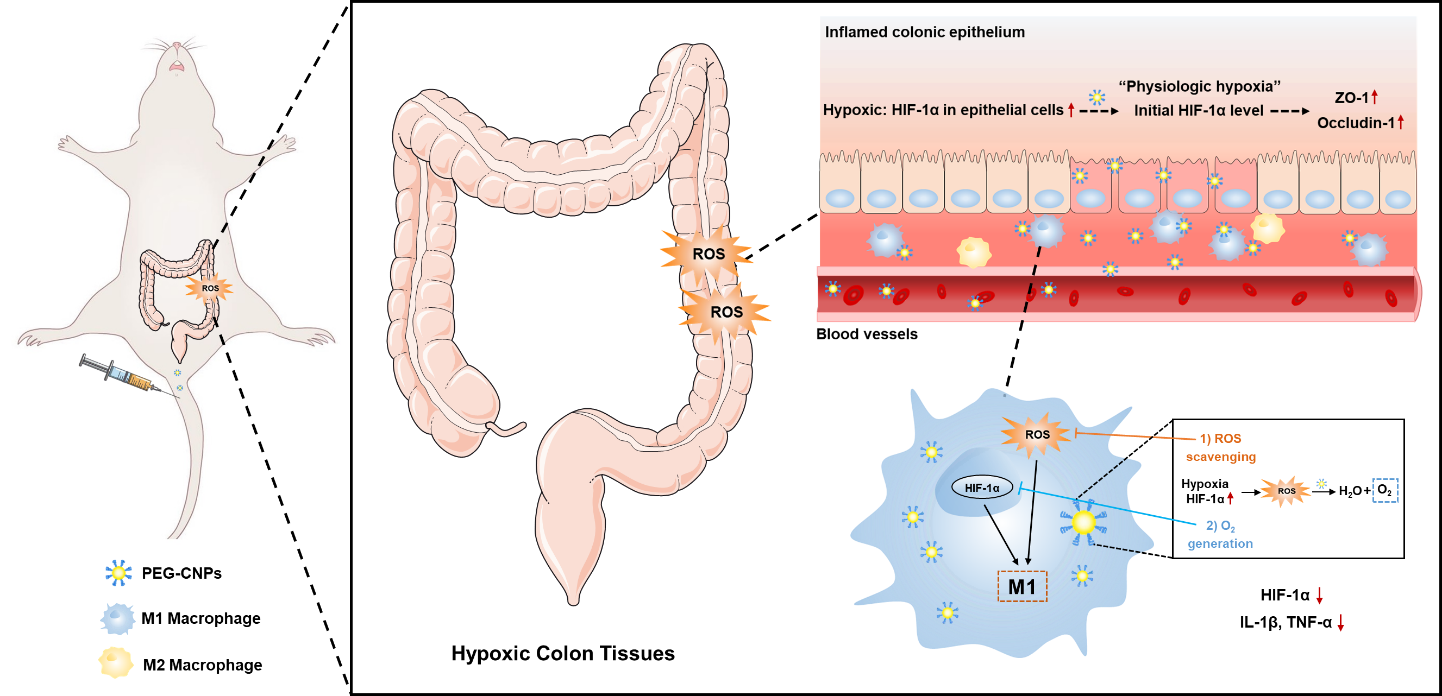
To investigate the protective effect of CNPs in colitis, a colitis mouse model was established by intracolonic TNBS administration to mice that fasted for 12 h. The mice were intravenously administrated with normal saline or 1.5 mg/kg CNPs at 2 h after colitis induction, and the therapeutic effect was evaluated 5 days later (Fig. 2a). In vivo colon-targeted analysis revealed a few CNPs in the colon of normal mice, while CNPs were distributed in the colons of colitis mice, indicating that CNPs preferentially accumulated at the inflammation site due to enhanced permeability and retention effect (EPR effect) (Fig. S7). This study evaluated the ameliorative effects of CNPs on TNBS-colitis using several indicators, including body weight, DAI, colon length, MPO activity and histological analysis. Mice in the TNBS group exhibited a decreasing trend in body weight, while there was a moderate increase in body weight in the CNPs group (Fig. 2b). The colon was significantly shorter in mice in the TNBS group (7.9 ± 0.4 cm) than in the control group (10.9 ± 0.4 cm) due to the colon inflammation, mucosal damage, and edema in the colon, while mice treated with CNPs had significantly improved colon lengths (9.7 ± 0.4 cm), nearly being restored to normal (Fig. 2d and 2e). Mice in the colitis group also exhibited a significant increase in DAI compared with those in the control group, while mice in the CNPs group had lower DAI than those in the colitis group (Fig. 2c). MPO, a glycosylase present in neutrophil and monocyte granules, can be acts as a biological marker to assess the disease status of UC [60]. Compared with the control group, MPO activity greatly increased in the colitis mice but significantly decreased in the CNPs treatment group, indicating CNPs reduced the infiltration of inflammatory cells in the colon (Fig. 2f). The histological analysis revealed that mice with colitis exhibited severe destruction of mucous membranes, massive inflammatory cell infiltration, goblet cell depletion, and disappearing crypt structures. In contrast, administration of CNPs significantly attenuated the histological signs of colitis, including tissue injury and inflammatory cell infiltration, indicating the significant protective effect of CNPs in colitis mice. These results indicate that CNPs can efficiently relieve the symptom of TNBS-induced colitis.
In vitro hypoxia attenuating and ROS scavenging of CNPs in intestinal epithelial cells
Previous studies have found that physiological HIF-1α expression provides a barrier-protective function in intestinal disease [61, 62]. We therefore next investigated the effects of CNPs treatment on HIF-1α expression in intestinal epithelial cells.
After 24 h of incubation using FITC-CNPs, cellular CNP uptake in HT-29 cells was confirmed using fluorescence microscopy (Fig. S8a). We verified the hypoxia-attenuating ability of CNPs by evaluating intracellular HIF-1α expression levels. HT-29 cells were incubated under hypoxia atmosphere (1% O2, 5% CO2, and 94% N2) and H2O2 for 4 h with CNPs treatment. As shown in Fig. 3a and 3b, we found that the initial HIF-1α expression in intestinal epithelial cells was comparable to that for “physiological hypoxia” under hypoxia condition, indicating that this condition could simulate the physiological hypoxia of intestinal epithelium. After adding H2O2, HIF-1α levels in HT-29 cells under hypoxia significantly increased. In contrast, HIF-1α levels significantly decreased to the initial level under hypoxia with CNPs treatment, which might be related to the O2 generation of CNPs during ROS scavenging.
Previous studies have found that stable HIF-1α expression in intestinal epithelial cells is helpful for repairing the intestinal mucosal barrier. Based on the hypoxia- attenuating ability of CNPs, we then explored the migration ability of HT-29 cells with CNPs treatment using a cell scratch test. As shown in Fig. 3c, HT-29 cells were cultured in a 12-well plate and scratched using a pipette tip to create scratches. After 24 h of incubation, marked migration of HT-29 cells was observed in the scratched area after CNPs treatment. The wound areas in the CNP group under hypoxia were significantly smaller than those in the other groups (Fig. 3c and 3d).
Considering the ROS scavenging capability of CNPs (Fig. 1d-f), we further evaluated the intracellular ROS levels in H2O2 treated HT-29 cells using a 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA) assay. As expected, a significant increase in ROS level was detected after H2O2 stimulation, which was markedly eradicated by the CNPs (Fig. 3e and 3f). To further confirm the anti-oxidative effect of CNPs in vitro, the cytoprotective effect of CNPs against H2O2 in HT-29 cells was determined. As illustrated in Fig. 3g, CNPs significantly decreased the H2O2-induced cell damage in a dose-dependent manner within 50 µM.
Nrf2 is known to be a major regulator of the cellular antioxidant pathways, which would be decreased due to excessive ROS. In response to oxidative stress, Nrf2 can dissociate from Keap1 and translocate into the nucleus, which plays a key role in initiating antioxidant response by activating a network of genes (such as Gpx1, Nqo1 and HO-1) [63, 64]. In this regard, we performed qRT-PCR analysis to determine the levels of Nrf2, DJ-1 and Keap1. The results indicated that the level of DJ-1 inhibited by oxidative stress could be reversed by CNPs (Fig. 4a). Meanwhile, CNPs upregulated Nrf-2 while downregulated Keap1, which contributed to the anti-oxidative ability of CNPs (Fig. 4b and 4c). To further investigate the role of Nrf2 in CNPs-mediated antioxidant activity, we examined whether Nrf2 silence could offset the protective effects of CNPs on H2O2-induced cytotoxicity (Fig. 4d and 4e). Studies have shown that the apoptosis of colonic epithelial cells can lead to the destruction of mucosal barrier and increase of inflammatory cells infiltration, and further aggravate mucosal injury [65]. Interestingly, CNPs would fail to prevent the HT-29 cells apoptosis after Nrf2 silence, suggesting that Nrf2 is a crucial factor in inhibiting apoptosis of CNPs (Fig. 4f and 4g). Furthermore, the status of DJ-1/Nrf2/Keap1 axis was analyzed by qRT-PCR analysis. Anti-oxidative stress genes (HO-1, Nqo1, Gpx1) up-regulation and pro-oxidative stress genes (Nox-2, Cyp2e1) in HT-29 cells treated with CNPs (Fig. 3h and 3i). In contrast, the relative levels of anti-oxidant genes and pro-oxidant genes in HT-29 cells with Nrf2 knockdown were no longer sensitive to CNPs treatment (Fig. 4h and 4i). These results indicated that CNPs were involved in activating Nrf2 to scavenge ROS (Fig. 4j), thereby preventing colonic epithelial cell apoptosis. In summary, the antioxidative activity of CNPs should be mediated by directly scavenging ROS and the reconstitution of the Nrf2-mediated cellular anti-oxidative system.
In vitro hypoxia attenuating, ROS scavenging and inflammatory regulation of macrophages induced by CNPs
Hypoxia of macrophages and the overproduction of ROS at the inflammation site of UC can aggravate the colon inflammation and induce the M1 polarization of macrophages [66]. Ameliorating hypoxia of macrophages and decreasing ROS levels can therefore attenuate pro-inflammatory macrophages, and effectively improve UC. To verify the hypoxia-attenuating ability of CNPs, we evaluated HIF-1α levels in vitro and intracellular oxygen levels. After 24 h of incubation with FITC-CNPs, fluorescence images illustrated the efficient cellular uptake of CNPs by RAW 264.7 cells (Fig. S8b). We also found up-regulation of HIF-1α expression in RAW 264.7 cells incubated with LPS for 24 h (Fig. 5a), which was consistent with previous studies [67]. We therefore carried out subsequent in vitro hypoxia-condition treatment with only LPS. RAW 264.7 cells were then incubated with LPS, and evaluated using [Ru(dpp)3]Cl2. [Ru(dpp)3]Cl2 is a typical indicator of intracellular oxygen which shows distinct red luminescence at 613 nm under hypoxia while the red luminescence is quenchable by oxygen [68]. As illustrated in Fig. 5b, compared with the LPS group, the treatment with CNPs led to significant decreases in red luminescence, indicating elevation of the intracellular O2 level. Subsequent Western blotting indicated that HIF-1α expression was significantly increased after LPS stimulation, which was attenuated by CNPs treatment (Fig. 5a).
Many macrophages are recruited and accumulated in the ulcerative colon, causing persistent inflammation. M1 macrophages produce pro-inflammatory cytokines and ROS [50]. To test the effect of CNPs on the inflammatory response in UC, the proportion of CD86+ cell was measured by flow cytometry. Interestingly, the proportion of CD86+ cells increased significantly due to the LPS stimulation, which could be reversed by CNPs treatment (Fig. 5c and 5d) and the morphological analysis of RAW 264.7 cells (Fig. S9). Moreover, pro-inflammatory M1 markers expression, including IL-1β and TNF-α, were significantly increased after LPS stimulation (Fig. 5e and 5f). Notably, treatment with CNPs markedly reduced the HIF-1α expression levels (Fig. 5g). A significant increase in the ROS level of RAW 264.7 cells was also detected after H2O2 stimulation, which was markedly attenuated by CNPs (Fig. 5h and 5i). HIF-1α, ROS, and M1 marker levels showed similar tendencies after the CNPs intervention, indicating that reducing HIF-1α expression and scavenging ROS play crucial roles in the suppression of M1 macrophages activation [57, 69]. Similarly, qRT-PCR analysis of RAW 264.7 cells showed that CNPs treatment significantly reduced the mRNA expression level of M1 markers including IL-1β and TNF-α (Fig. S10). Together these observations indicated that CNPs can attenuate hypoxia of macrophages and scavenge excess ROS in macrophages via generate O2 and scavenging ROS, and the successful inhibition of M1 macrophages activation.
Restoration of colonic epithelium and inhibition of M1 macrophages activation induced by CNPs in the colitis mice
To further elucidate the mechanism of the protective effect of UC, we investigated the impact of CNPs on colonic epithelium restoration and colonic M1 macrophage in colitis mice. We first isolated colon tissues to track the intracellular fate of FITC-CNPs at 0.5 h after the injection. Confocal laser scanning microscopy images of colon tissues illustrated that FITC-CNPs were distributed more in the inflammatory cell infiltration layer of colon tissues than in the colonic epithelium (Fig. 6a). We then determined whether O2 generated by CNPs alleviated hypoxia in inflamed colons. Immunohistochemical HIF-1α staining demonstrated that CNPs administration prominently inhibited HIF-1α expression in the inflammatory cell infiltration layer of colon tissues (Fig. 6b).
We investigated the effect of CNPs on colonic epithelium with disrupted intestinal barrier functions caused by TNBS. The effects of HIF-1α protein stabilization on barrier protection are thought to be attributable to tight-junction protein (such as claudin-1) regulation [70, 71]. In the present study, colitis mice administered with CNPs had normalized ZO-1 and occludin-1 levels, which are tight-junction proteins that play vital roles in gut homeostasis (Fig. 6d-f) [3]. Furthermore, compared with the TNBS group, CNPs prevented systemic FITC-dextran exposure in colitis mice (Fig. 6c), demonstrating intestinal barrier function restoration. Collectively these results indicated that CNPs could enhance HIF-1α protein stability in intestinal epithelial cells and restore intestinal barrier functions by increasing the expression of tight-junction proteins [72].
The role of CNPs in suppressing the pro-inflammatory macrophage activation in vivo, the levels of CD86+ cells in the colon were measured immunohistochemically. As shown in Fig. 7e and 7f, the CD86 level was increased after inducing colitis, whereas CNPs injection prominently reduced that level. Besides, consistent with the in vitro results (Fig. 5e and 5f), pro-inflammatory cytokine levels (including IL-1β and TNF-α) in the colon tissues and blood serum were lower in the CNPs group than in the colitis group (Fig. 7a-7d), suggesting that CNPs suppress inflammation. To further examine the anti-oxidative effect of CNPs in vivo, we measured MDA levels and SOD activity in colon tissues. The MDA level was elevated and SOD activity was inhibited after TNBS treatment, while treatment with CNPs could reverse the imbalance of these ROS-related indicators (Fig. 7g and 7h). Consequently, CNPs suppress pro-inflammatory macrophage activation by alleviating hypoxia and scavenging ROS of colon inflammatory tissues, while enhancing the body’s ability of ROS scavenging.
{kind=link}