We have demonstrated the potential of using mitochondrial 12S and 16S rRNA genes as genetic markers for molecular systematics of parasitic nematodes through comparisons with other commonly used genetic markers. We have also evaluated their utility through phylum-wide phylogenetic analyzes, providing evidence of their value as robust genetic markers for the study of nematode relationships.
Potential utility of mitochondrial 12S rRNA gene in nematode molecular systematics
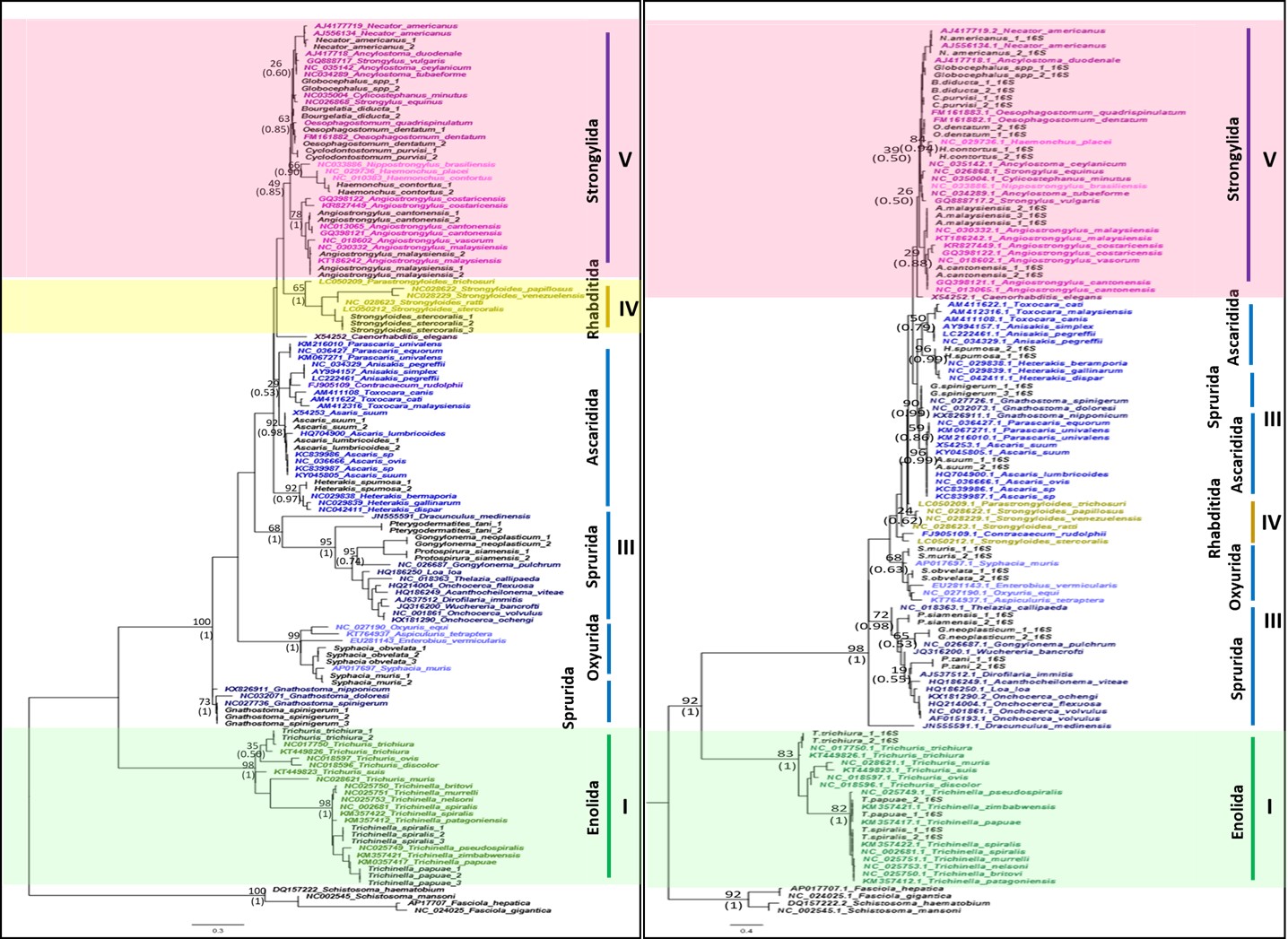
Phylogenetic relationships were inferred using 120 sequences from 75 species of nematodes, showing the potential of the 12S rRNA gene as an effective marker for nematode molecular systematics. First, clades I, IV, and V were monophyletic in our analyzes, revealing that this genetic marker has sufficient resolution to discriminate between the clades. Although clade III being not monophyletic, our results showed a congruence with previous studies that used whole mitochondrial genomes, in which clade III was not monophyletic [8–10]. The non-monophyly of clade III, consisting of orders Ascaridida, Spirurida, and Oxyurida, remains unresolved. However, this demonstrates that the 12S rRNA gene is robust for molecular systematics, provides sufficient resolution, and is comparable to the whole mitochondrial genome. Even though the partial 12S and 16S rRNA sequences are only 460 bp and 240 bp in length, respectively, amplification of a short-length genetic target to study nematode phylogeny is undoubtedly more useful and practical than sequencing whole genomes. For example, long-preserved specimens which may be affected by genetic degeneration issue (e.g., DNA template shearing or fragmentation), could get benefit from short-length genetic amplification.
Second, mitochondrial 12S rRNA showed that there was a close relationship between clades IV and V, which has also been found in studies using the 18S rRNA gene and the transcriptome [7, 11–12, 30]. However, compared to phylogenetic studies using whole mitochondrial genomes, clade V was more closely related to Ascaridida, whereas clade IV was a sister clade to the clade consisting of clade V and Ascaridida [8–10]. Strongyloides and Parastrongyloides, the clade IV representative species used in this study, have conventionally been classified to the order Rhabditida, with a close relationship to clade V nematodes. However, they seem to be distantly related to other members of clade IV based on morphology. However, studies using molecular techniques have revealed a different phylogenetic position for Strongyloides. Using whole mitochondrial genomes, Lv et al. revealed that Strongyloides is nested within clade III nematodes, and the authors also suggested that clade IV may be paraphyletic [31].
Third, because the 12S rRNA gene showed clade IV to be monophyletic, it has greater discriminatory potential than 16S rRNA. The 12S rRNA gene provided sufficient discrimination between clades in terms of molecular systematics for nematodes, with monophyly found for three of the four clades. Even though the tree topology from concatenated 12S and 16S rRNA gene analysis also resulted in monophyly for three of the four clades, sufficient resolution was gained from the use of a single gene; thus, the use of concatenated genes is unnecessary. Lastly, the potential benefit of the 12S rRNA gene as a genetic marker was highlighted by its amplification efficacy, able to amplify from a single larva. The sensitivity and specificity of this genetic marker indicate that it is a convenient and robust method for the study of molecular systematics. Taken together, the advantages of utilizing the 12S rRNA gene as a genetic marker for molecular systematics of nematodes were demonstrated through the ease of its application and its congruence with phylogenies from whole mitochondrial genomes, as well as from the nuclear 18S rRNA gene.
Non-monophyly of clade III nematodes and phylogenetic position ofGnathostoma
Clade III, containing orders Ascaridida, Spirurida, and Oxyurida, was not found to be monophyletic in any of our analyzes. In current studies of nematode phylogenies, mitochondrial genes have shown the nematode clade III not to be monophyletic, whereas, the working hypothesis of 18S rRNA gene phylogenies is that clade III is monophyletic. However, Nadler et al. showed that clade III was not monophyletic by using the 18S rRNA gene due to the inclusion of the superfamily Gnathostomatoidea [32]. Phylogenies from another study that used the 18S rRNA gene focused on nematodes belonging to Spirurida; in addition, the Gnathostomatoidea were not placed with the rest of Spirurida but instead formed a clade at the basal position of the phylogenetic tree [33]. Similarly, Liu et al. could not resolve the phylogenetic position of Gnathostoma using mitochondrial genomes [34]. The authors used the complete mitochondrial genomes of 58 species of nematodes and found that Gnathostoma was closely related to Ascaridida but was not placed with the rest of Spirurida. Despite many studies and the use of various genetic markers attempting to resolve these discrepancies, the phylogenetic position of clade III nematodes remains inconclusive. A possible explanation is that the phylogenetic position of Gnathostoma is based on the traditional classification using morphological characteristics. Gnathostoma belongs to order Spirurida and is the typical genus of the superfamily Gnathostomatoidea. Nematodes belonging to Spirurida have a bilaterally symmetrical anterior extremity, the lateral external labial papillae are absent, and they mostly dwell in the tissues of the host [5, 6]. However, when the phylogenetic position of Gnathostoma was determined based on molecular genetic markers, an incongruence between morphological and molecular phylogenies was revealed. We propose the inclusion of Gnathostoma into future nematode phylogenies for a more accurate picture of its taxonomy. A more comprehensive study with a focus on only clade III nematodes or the inclusion of a greater number of taxa may provide more clarity of nematode phylogenies.
Advantage of mitochondrial 12S and 16S rRNA over nuclear 18S rRNA gene
The current working hypothesis, first proposed by Blaxter et al., for the molecular systematics of nematodes used 18S rRNA gene-based phylogenies [7]. However, recently, mitochondrial genomes have served as alternative genetic markers for determining nematode relationships [8–10]. Mitochondrial DNA has a faster evolutionary rate compared to nuclear ribosomal genes such as 18S rRNA, and this confers an advantage to mitochondrial genes because of their ability to discriminate closely related species [13–15]. Several studies using the nuclear 18S rRNA gene were unable to provide taxonomic clarity at the species level, as there was insufficient sequence variation between species. In the closely related and morphologically similar Haemonchus placei and H. contortus, the nuclear 18S rRNA gene sequences are identical [35–37]. Among Trichuris spp., morphological variations also confound morphological identification [38, 39]. Moreover, the highly similar nuclear 18S rRNA sequences of T. vulpis and T. serrata indicate that this gene might not have sufficient sequence variation to discriminate between closely related species [39].
In this study, we demonstrated the resolution power of mitochondrial 12S and 16S rRNA genes for species discrimination through the phylogenetic placements of nematode species using these two genetic markers. Despite the nuclear 18S rRNA gene provides better resolution at higher taxonomic levels because of the sufficient variation between clades, as seen in monophyletic clades, the gene’s resolution at lower taxonomic levels remains ambiguous, as seen in the species discrimination results. Undoubtedly, mitochondrial 12S and 16S rRNA genes have advantages over nuclear 18S rRNA as phylogenetic markers. They provided a better resolution at lower taxonomic levels because of sufficient genetic variation between species, as suggested by the phylogenetic placement of nematode species in our analyzes, as well as a resolution at higher taxonomic levels as a result of sufficient genetic variation between clades, except for clade III, as indicated by monophyly. Thus, utilizing mitochondrial 12S and 16S rRNA genes as genetic markers for nematode molecular systematics may not only provide resolution at lower taxonomic levels but also higher levels across a wide-ranging taxonomic hierarchy.
Potential use of mitochondrial 12S and 16S rRNA genes for nematode DNA barcoding
A DNA barcode is defined as a short sequence of DNA that can accurately identify organisms [40–42]. The concept of DNA barcoding was first proposed by Herbert and colleagues when they suggested using DNA sequences as taxon ‘barcodes’ for the global identification of all organisms [42]. Species are usually identified by comparing unknown sequences against known reference sequences. Currently, the universal genetic marker used for DNA barcoding is the 5ʹ end of the mitochondrial COI gene. The COI gene has specific properties that make it useful as a genetic marker for DNA barcoding, including multiple copies, robust PCR amplification, nearly identical sequences in individuals of the same species yet sufficient variation between species, and the presence of a reference database for sequence comparison [40–42]. In this study, the viability of using mitochondrial 12S and 16S rRNA genes for DNA barcode identification of nematodes species was demonstrated; 100% of our representative nematodes species were accurately taxonomically assigned, suggesting that both genes have sufficient genetic variation to allow for species identification. Furthermore, as both 12S and 16S rRNA genes are mitochondrial in origin, PCR amplification success and robustness will be comparable to the COI gene. The mitochondrial 12S and 16S rRNA genes contain more highly conserved sequences than COI, facilitating easier primer design, but also contain sufficient genetic variation for species identification.
Although primers had to be designed separately for nematodes of clade I and nematodes of clades III, IV, and V, the same region was used for primer design. This enabled DNA sequence and phylogenetic analyzes of the clades to be performed together, maximizing the range of taxa studied. Although the current universal gene for DNA barcoding studies of animals is COI, an alternative gene for species identification could be useful. Studies have evaluated the potential use of the mitochondrial 16S rRNA gene as a DNA barcode for insects [43, 44]. A comparison between mitochondrial COI and 16S rRNA gene was performed, and the authors concluded that the 16S rRNA gene could detect more taxa than COI because of reduced PCR amplification bias.
Ferri et al. compared the performance of mitochondrial COI and 12S rRNA genes for DNA barcoding of Spirurida nematodes and suggested that the 12S rRNA gene could be appropriate for species-level identification [45]. However, in order for mitochondrial 12S and 16S rRNA genes to be utilized in DNA barcoding, it is essential to establish sufficient sequences in the reference databases. The advantage of the COI gene over other genetic markers, such as mitochondrial 12S and 16S rRNA genes, is the extensive availability of database sequences, allowing for thorough comparisons of unknown sequences. However, the potential of mitochondrial 12S and 16S rRNA genes for DNA barcoding of nematodes cannot be disregarded, as we have shown the capability of these two genetic markers to discriminate between species. Therefore, mitochondrial ribosomal genes have considerable value, not only as markers for molecular systematics studies but also their potential use in DNA barcoding studies. Future studies are needed to evaluate further the potential and utility of mitochondrial rRNA genes for DNA barcoding and to expand the reference databases for these genetic markers.
Limitations
Clade II and plant-parasitic nematodes belonging to clade IV were not included in this study. The exclusion of representative species from these two groups could have resulted in an incomplete picture of phylum-wide relationships. Moreover, the primers designed in this study are clade-specific because it was challenging to find regions conserved across the four clades. Primers had to be designed separately for nematodes belonging to clade I and nematodes belonging to clades III, IV, and V. An increase in the number of taxa could potentially aid in the generation of a complete picture of phylum-wide relationships.
{kind=link}