We performed a comprehensive representation analysis to study the association between Ts1Cje Differentially Expressed Genes (DEGs) and mouse REST target genes. This analysis encompassed the microarray datasets from postnatal, embryonic, and neural progenitor stages. We found that Ts1Cje DEGs significantly overlapped with the mouse REST target genes and were over-represented at most developmental stages except embryonic UpDEGs is under-represented (representation factor = 0.8). This indicates that REST dysregulation may be the culprit for the DS complex clinical and neuropathology development. Particularly in the neural progenitor stage dataset, the DEGs are 2-fold over-represented with REST target genes. This indicates REST plays an essential role in this stage, and their dysregulation in DS contributed to complicated neuropathology.
Database for Annotation, Visualisation, and Integrated Discovery (DAVID)(Sherman et al., 2022) was used to perform the Gene Ontology (GO) enrichment analysis for the overlapped genes and revealed the biological processes involved by these significantly up- or downregulated DEGs which are targeted by REST at the same time. In the postnatal stage, the overlapped UpDEGs and DownDEGs with REST target genes are involved in the interferon-related and cell cycle-related processes respectively. For UpDEGs, the most significant GO is the defence response to the virus (GO:0051607) with p-value < 0.001 as the main function of interferon in the body's immune system to combat the virus infection and the other top 10 GOs are involved in the interferon regulation and production. Interferons (IFNs) are cytokines involved in the body's innate immune system to protect against viral infections and inflammation conditions (de Andrea et al., 2002; Kopitar-Jerala, 2017). The enrichment results show that the genes encoding the production of Type 1 IFNs (including IFN-alpha, IFN-beta, IFN-kappa, and IFN-omega) and their specific cell surface receptor IFN-α/β (IFNAR) are significantly upregulated. Previous publications proposed that IFN hyperactivity and autoimmunity are commonly seen in DS individuals (Araya et al., 2019; Jagadeesh et al., 2020; Malle & Bogunovic, 2021). These observations agree with the gene dosage imbalance as four subunits of IFN receptor genes are located on human chromosome 21 (HSA21), and trisomy in Down syndrome causes their over-expression. Interestingly, we found that these IFN genes are also targeted by REST which suggests that the REST dysregulation also contributes to the IFN's upregulation in DS (Chung et al., 2021; Kirsammer & Crispino, 2016). Furthermore, the upregulated IFNs will over-activate the downstream IFN-stimulated genes (ISGs) and promote inflammatory and antiviral activity (Jagadeesh et al., 2020). For instance, over-activation of JAK-STAT which is one of the pro-inflammatory pathways, and it was proposed that the JAK-STAT signalling pathway is involved in the DS neuropathology, including the neurogenic-to-gliogenic shift (Banerjee et al., 2017; Lee et al., 2016; Malle & Bogunovic, 2021).
On the other hand, the postnatal DownDEGs overlapped with mouse REST are significantly grouped into the cell cycle-related processes, including cell division, DNA replication, and various cell cycle regulators. Downregulation of these genes is aligned with the notion that neurogenesis impairment is one of the preserved neuropathology seen in various DS models and causes cognitive impairment (Hwang & Zukin, 2018; Lott & Dierssen, 2010). Previous studies supported that the altered cell cycle can cause reduced cell proliferation which starts from the early embryonic to the postnatal development in various DS mouse models and human induced pluripotent stem cells (Contestabile et al., 2007; Ishihara, 2021; Stagni et al., 2018; Tang et al., 2021; Utagawa et al., 2022). Therefore, this GO analysis revealed that these cell cycle genes were targeted by REST and their downregulation in DS may be attributed to the REST dysregulation.
In the embryonic stage, the UpDEGs are related to cell regulation and cell death processes. The genome-wide gene dysregulation is always seen in DS models, and the imbalanced expression of these genes can disturb the developmental pathways and be associated with DS pathological development (Letourneau et al., 2014; Lockstone et al., 2007). The trisomy effect of HSA21 has been proposed to be the main factor that causes genome-wide gene dysregulation, including the "amplified developmental instability theory", "gene dosage effect" and "Down syndrome critical region" theories (Amano et al., 2004; Korenberg et al., 1994; Shapiro, 1975). Intriguingly, this study further adds that REST as an epigenetic gene repressor which significantly targets these cell regulation genes thus potentially contribute to genome-wide gene dysregulation and leading to DS neuropathology. Moreover, the UpDEGs involved in cell death processes are aligned with previous studies which proposed the excessive developmental apoptosis (programmed cell death) in DS associated with neurodegeneration and cognitive impairment (Rueda et al., 2013; Seidl et al., 2001). On the other hand, the DownDEGs are associated with endocytosis, lipid metabolism, cell cycle nucleotide and signalling molecules biosynthesis, and various molecule modification processes, including phosphorylation, protein phosphorylation, and methylation. These downregulated processes may disrupt embryonic brain development, especially synaptic formation. Lipid metabolic and transport processes are essential in synapse development and maintenance (Lockstone et al., 2007). Previous studies proposed that the availability of cholesterol is the bottleneck limit for synaptogenesis as they play a crucial role in the functional regulation of synaptic vesicles exo- and endocytosis also for the biosynthesis of signalling molecules and neurotransmitters (Pfrieger, 2003a, 2003b).
In the neural progenitor stage, the neurospheres show both REST targeted UpDEGs and DownDEGs are significantly involved in the cell cycle-related processes and DNA repair processes. Neural progenitor cells are actively proliferating and differentiating into neural cells at this stage. These GOs suggest that specific brain cell populations were up- and certain were downregulated in the DS neural cell population. This disproportionate brain cell population may exacerbate the effectiveness of functional neurons to form connections and transmit impulses, causing DS neuropathology for instance cognitive and memory impairments. In addition, the REST-targeted DownDEGs involved in protein synthesis suggest that the proliferated cells may not have functional protein expression for neural differentiation and contribute to DS neuropathology.
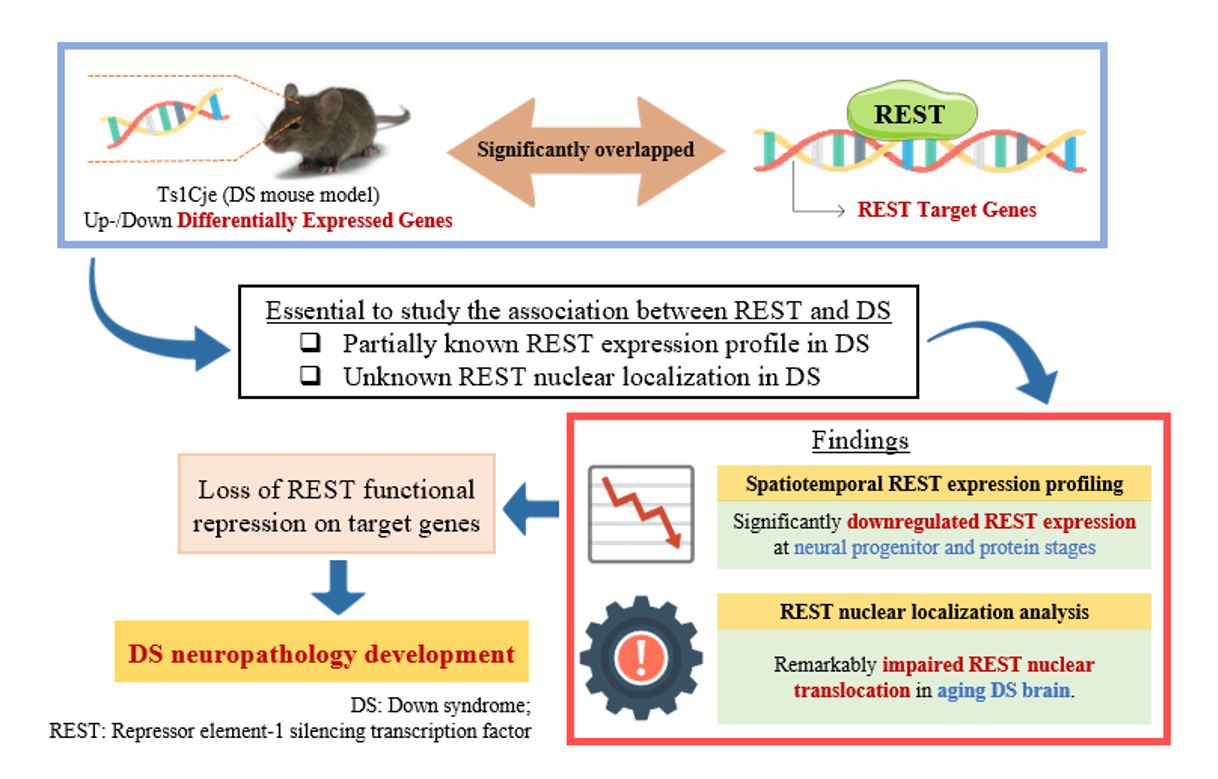
In short, we performed a comprehensive representation analysis based on the DEGs of Ts1Cje DS mouse model at postnatal, embryonic, and neural progenitor stages. We revealed that these DEGs overlap with mouse REST targets, emphasising the crucial role of REST expression and functional repression in DS. Hence, the characterisation of spatiotemporal Rest expression and REST nuclear translocation is essential to study the association between REST and DS for understanding how the REST contributes to the Up-and DownDEGs, which causes the DS neuropathology.
The spatiotemporal RT-qPCR analysis shows that the Rest expression is not significantly different between Ts1Cje and WT models at most postnatal stages. The cerebral cortex, especially the prefrontal cortex, is the most vulnerable brain region to the negative effect of stress and uncontrollable stress can result in the rapid loss of cognitive function (Arnsten, 2009). Our results show a significant spike in Rest expression at Postnatal Day-30 is aligned with this situation as REST is involved in stress resilience, especially with higher stress in the adult stage (Mampay & Sheridan, 2019). In addition, DS adults were proposed to associate with oxidative stresses and caused memory declination, cognitive impairment, and early onset of Alzheimer's disease (Perluigi & Butterfield, 2012; Strydom et al., 2009; Zis et al., 2012). Hence, the increased Rest expression at P30 suggests the REST function in regulating neuroprotection and stress resilience.
In the hippocampus, the significantly downregulated Rest expression at P84 in the Ts1Cje model suggests the greater loss of REST neuroprotective function in DS individuals at the later stage of life. Previous publications proposed that 15–77% of DS individuals over 60 years are affected by dementia and Alzheimer's disease. It is commonly diagnosed between the fourth to sixth decades of DS individual life with a significant deposited amyloid plaque and neurofibrillary tangles starting in the hippocampus (Head et al., 2012; Lott & Dierssen, 2010; Nelson et al., 2011). On the other perspective, Rest was proposed to be upregulated in WT ageing brains for neuroprotection, suppression of β-amyloid toxicity, and oxidative stress-related genes to preserve neuronal viability and cognitive functions (Hwang & Zukin, 2018; Lu et al., 2014). Hence, loss of Rest in DS individuals may lead to a deficiency in neuroprotection and stress resilience that can cause neurodegeneration.
In the prenatal stages, the differences in Rest expression between both DS and WT samples were hypothesised as they were proposed to have higher Rest expression for functional repression of the neuronal genes in embryonic stem cells (ESCs) and neuronal stem cells before neurogenesis (Jørgensen et al., 2009). However, we found that Rest expression is not significantly different between Ts1Cje and WT models at the embryonic stage. We did not see a significant difference in Rest expression at the temporal analysis across the prenatal E14 and postnatal P1.5 timepoints, suggesting that Rest expression is maintained for both mouse models before and after birth.
Neurospheres are a mixed population of neural stem cells, including neural and glial progenitor cells. This in vitro model allows the study of Rest expression in the early neurodevelopmental stages and contains a more homogeneous neural progenitor cell population compared to the whole/regional brain samples used in the postnatal and embryonic stages. Interestingly, the neurosphere samples show a strongly significant difference in Rest expression between Ts1Cje and wildtype mice which indicates REST dysregulation happens earlier in the progenitor stage. Although our postnatal and embryonic results show Rest expression is restored to wildtype level, the DS neuropathology suggests that Rest dysregulation happened in the neural progenitor stage brings irreversible pathological effects. Furthermore, these results also suggest that Rest expression may be masked by the heterogeneous brain cell population in the whole/regional brain samples at the postnatal and embryonic stages as different brain cell types express different Rest expression levels. Additionally, the neurogenic-to-gliogenic shift proposed in DS with more glial cells present from the early neurodevelopment stage may compensate for and mask the defective target neuron Rest expression in the whole/regional brain samples (Lee et al., 2016). This is a novel insight regarding the heterogeneous brain cell types that may influence the Rest expression study and warrant further study.
To summarise, RT-qPCR results show that the Rest expression is significantly downregulated in the Ts1Cje mouse model against wildtype at the more homogeneous neural progenitor stage. This result concurs with previous studies which proposed that Rest downregulation in early developmental stages accelerated premature neural differentiation and depleted the neural stem cell pool (Canzonetta et al., 2008). The pre-matured neurons were found to malfunction (Hanson et al., 2007; Kaufmann & Moser, 2000), and deficient neural stem cells turned over to replace lost neurons, which led to various DS neuropathology and cognitive impairment. In addition, Rest downregulation in the neural progenitor stage may also contribute to the disrupted early cell-fate determination, which is one of the DS neuropathology that neural progenitor cells were proposed to shift into the glial lineages (known as neurogenic-to-gliogenic shift). Janus kinase-signal transducer and activator of transcription (JAK-STAT) is a well-studied gliogenic signalling pathway and will be activated upon the neuronal-to-glial differentiation transition (Lee et al., 2016; Ling et al., 2014). This signalling pathway was proposed to be dysregulated in DS to explain the neurogenic-to-gliogenic shift (Bonni et al., 1997; Hewitt et al., 2010; Lee et al., 2016, 2019). Surprisingly, this JAK-STAT signaling pathway and their upstream activators, the interferon signalling cascade, were also targeted by REST according to the collection of processed chromatin immunoprecipitation sequencing (Rouillard et al., 2016) and our bioinformatics enrichment analysis in the previous section.
In the Western blot study, it was demonstrated that REST expression is significantly different between the Ts1Cje and wildtype models. The expression fold change of REST in DS is 2-fold-downregulated against WT. This has proven that REST can be highly expressed in a normal healthy ageing brain to exert neuroprotection against stressors (Lu et al., 2014). In contrast, being at high risk of having early onset Alzheimer's disease (Lautarescu et al., 2017; Salehi et al., 2015), this neuroprotection is diminished in DS brains since REST is significantly downregulated in DS individuals. Therefore, it can be suggested that REST is a potential candidate for the therapeutic target to improve intellectual and cognitive behaviour among DS individuals.
Impairment of REST protein nuclear translocation is well-described in neurodegenerative diseases including Alzheimer's disease, Parkinson's disease, and Huntington's disease but has not been described in Down syndrome yet (Kawamura et al., 2019; Lu et al., 2014; Shimojo, 2008; Zuccato et al., 2007). Hence, this study is the first to reveal the association between REST nuclear localisation and Down syndrome. This perinuclear marginalisation effect of the Ts1Cje model is also seen in previous publications which indicate the impairment of REST nuclear translocation and loss of REST neuroprotection (Kawamura et al., 2019; Lu et al., 2014). REST binding epigenetically and repressing the target gene's function, the impairment of nuclear translocation in this study suggests that REST has lost the functional neuroprotection and stress resilience in the ageing stage. This supports the notion that neuroinflammation and neurodegeneration diseases are commonly diagnosed in DS individuals at the later stage of life (Lu et al., 2014). REST epigenetically targets more than half of human genes and for instance, previously proposed inflammatory-associated genes trisomy in DS including the interferon receptor genes IFNAR1, IFNAR2, IFNGR1 & IFNGR2 (Wilcock, 2012). The failure of REST nuclear translocation may attribute to the over-expression of REST target inflammatory-associated genes and lead to uncontrolled inflammation and neurodegeneration. In alignment with our qPCR results in the postnatal stages hippocampus region, the loss of REST expression along the developmental timepoints and impaired REST nuclear translocation at the ageing stage may contribute to the commonly developed early-onset of Alzheimer's disease in DS individuals. In short, this study reveals that the impairment of REST nuclear translocation in the ageing stage of the DS mouse model and the loss of REST functional repression for neuroprotection & stress resilience may contribute to DS neuropathology.
{kind=link}