We herein explored the processes shaping bacterial communities within the phycosphere in response to phytoplanktonic extracellular fractions (PEF) by considering growth patterns and community assemblages. We first assembled bacterial isolates, comparable to those found associated with phytoplankton in the environment, into a synthetic community (SynCom) and used the PEF as substrates to test their impact on the growth of both the bacterial isolates and SynCom. In a second experiment, the SynCom was co-cultivated in the presence of live algal cells. The community composition was examined in both experiments to test whether the presence or absence of algae may influence the bacterial assemblage. That way, we developed an experimental approach to gradually take the biological complexity of the algal/bacterial system into consideration, from PEF exposure on bacterial isolates to a more complex microbial interaction, involving algae/bacteria and bacteria/bacteria interactions.
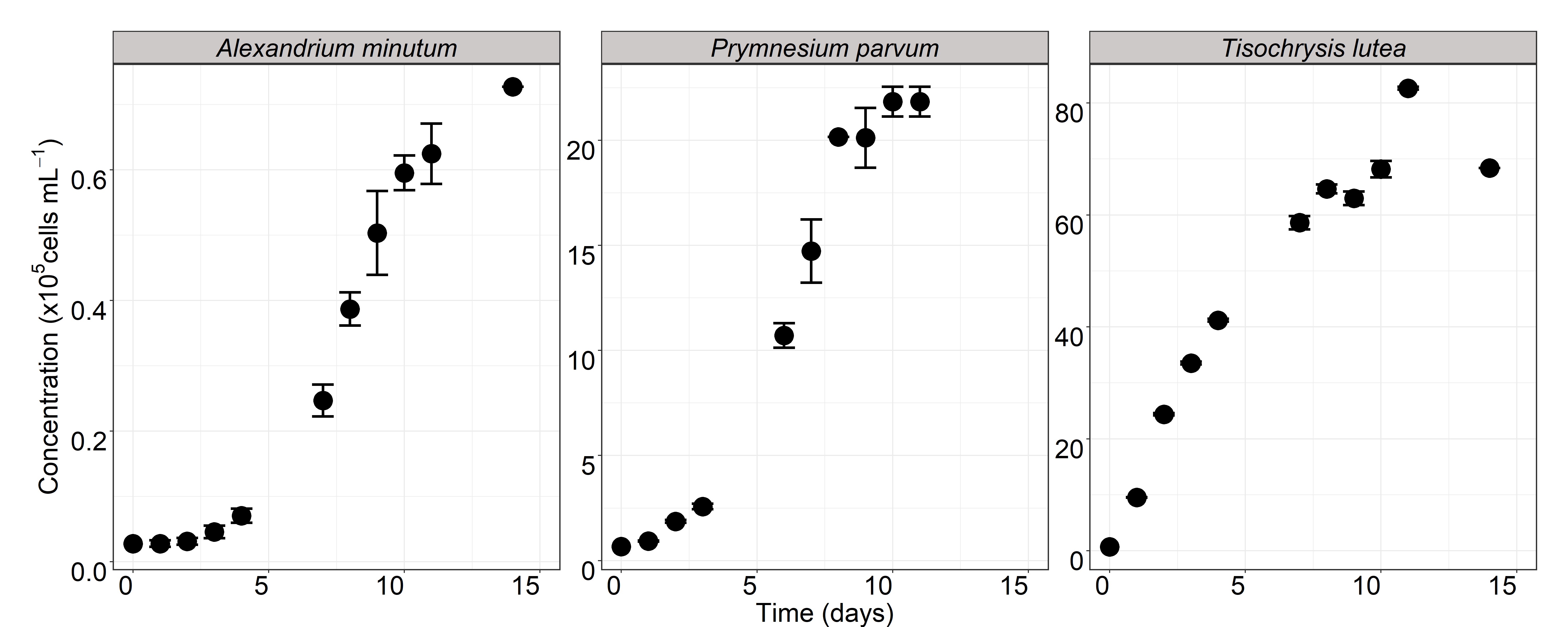
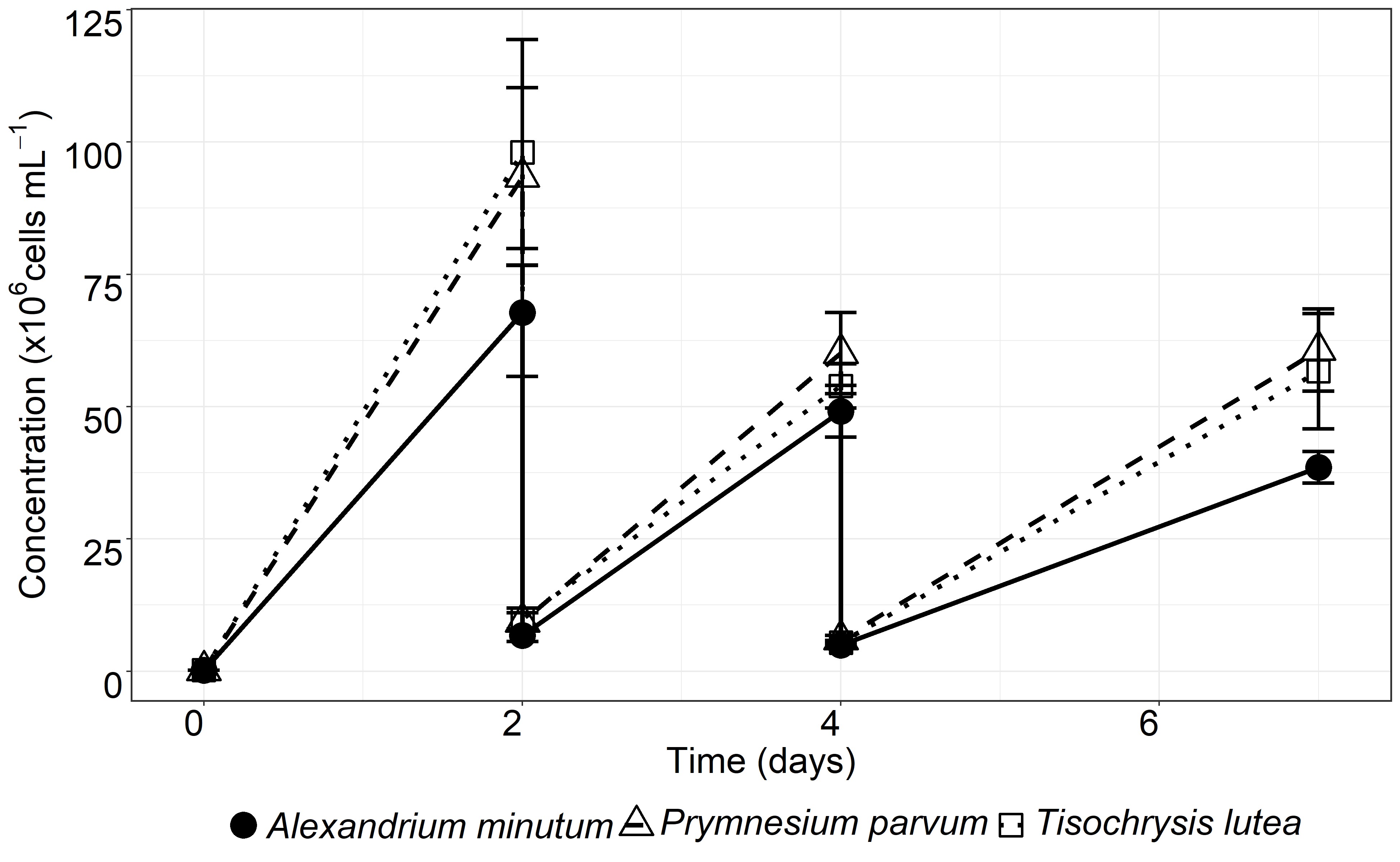
The PEF analysis of the three phytoplanktonic species (A. minutum, P. parvum and T. lutea) revealed high inorganic N/P ratios, suggesting a global P deficiency. The P was mainly sequestered in phytoplankton cells during P-limited axenic algal cultures used for PEF preparation. Here, the sole addition of PEF to bacterial isolates (except for P. parvum) and SynCom (except for T. lutea) increased bacterial abundance compared to SW + CMM, consistently confirming that algal PEF provided additional compounds (i.e. organic nutrients) that a complete mineral medium did not. In classical growth media, the bacterial growth yield depends on the initial concentration of the limiting nutrient (67). The addition of CMM in PEF lifted an additional level of limitation in mineral elements, allowing reaching an even higher bacterial cell abundance in the isolates and SynCom. The addition to the PEF of P alone showed a much stronger effect on the SynCom bacterial abundance than the sole addition of N. These observations supported the common assertion that heterotrophic bacteria use the DOM secreted by microalgae and confirmed that the limiting element, phosphorus here, was the trigger of bacterial growth.
Bacterial growth inhibition was never observed (Fig. 1), suggesting the absence of antibacterial compounds in the PEF. Substantial differences of final concentrations were noted between isolates that may be related to morphological differences between bacteria resulting in differences in cell volumes and total cell contents (68). The growth of both individual bacteria and SynCom led to the same concentration range in PEF + CMM cultures, suggesting that the bacteria must share trophic resources in the SynCom. Competition for trophic resources can involve many aspects such as uptake kinetics, growth rates and direct interaction between cells (see for review (69)). In the present study, we acknowledge that the description of the bacterial growth was based on every two days record in fed batch cultures and did not give information on the maximal growth rates of each bacterial isolate, but rather on their ability to reach a concentration in two days. Therefore, potential differences in maximal growth rates between strains, that may give a competitive advantage, cannot be ruled out. Some algal exometabolites are known to partially affect bacterial growth (70) and among them exometabolites from P. parvum can inhibit leucine incorporation by bacteria (71). This could explain the absence of isolate growth in PEFP. parvum but this hypothesis seems unlikely given the observed growth of isolates on PEFP. parvum+CMM and that of SynCom on PEFP. parvum and PEFP. parvum+CMM. Conversely, all isolates, but not SynCom, were able to grow on PEFT. lutea. Limited access to phosphorus or microbial interaction processes may be involved but require further investigation. Of note, in the co-culture experiment, algal growth was never impaired by the presence of bacteria, indicating the absence of algicidal compounds.
The specificity of bacterial communities in phycosphere and the mechanisms involved in species assembly are still largely unknown. In general, the phycosphere is mainly dominated by heterotrophic bacteria belonging to Bacteroidia, Alpha- and Gamma-proteobacteria (8). However, molecular analyses reveal a wide diversity inside these groups and a specific composition related to numerous parameters including algal species and ecotype, ecophysiological conditions and sampling location. For instance, the intraspecific genotype seems to be the main driver of microbiomes of the diatom Thalassiospira rotula (26), whereas location and time from which the diatom Leptocylindrus is isolated play important roles in shaping its microbiome (72). Here, the use of a SynCom limited the influence of host lineage, geographical isolation and strain maintenance conditions. We first investigated how natural phytoplankton-derived extracellular compounds modulated SynCom assembly without the contribution of living phytoplankton cells; and explored in a second time the underlying mechanisms in a more complex biological system by co-cultivating SynCom with each of the two harmful microalgae.
Our metabarcoding analyses revealed that whatever the algae, the most abundant bacteria in PEF in both free-living and attached fractions were Alteromonas. In a recent study, using a miniaturized co-culture setup directly in the field, Raina et al. (73) investigated the capacity of distinct phytoplankton-derived DOM to attract specific bacteria and archaea assemblages. Their results showed that DOM-derived from A. minutum and P. parvum were the strongest chemoattractants amongst the ten phytoplankton lysates tested. These results evidenced that these species are chemical hotspots enabling the enrichment of the phycosphere with copiotroph taxa like Alteromonas spp. This ubiquitous marine Gammaproteobacteria is indeed highly abundant in enriched DOM environments (74–76), including phytoplankton cultures and blooms (8, 77, 78), where they easily thrive due to their hydrolytic capacities of diverse algal-derived polysaccharides (79–81).
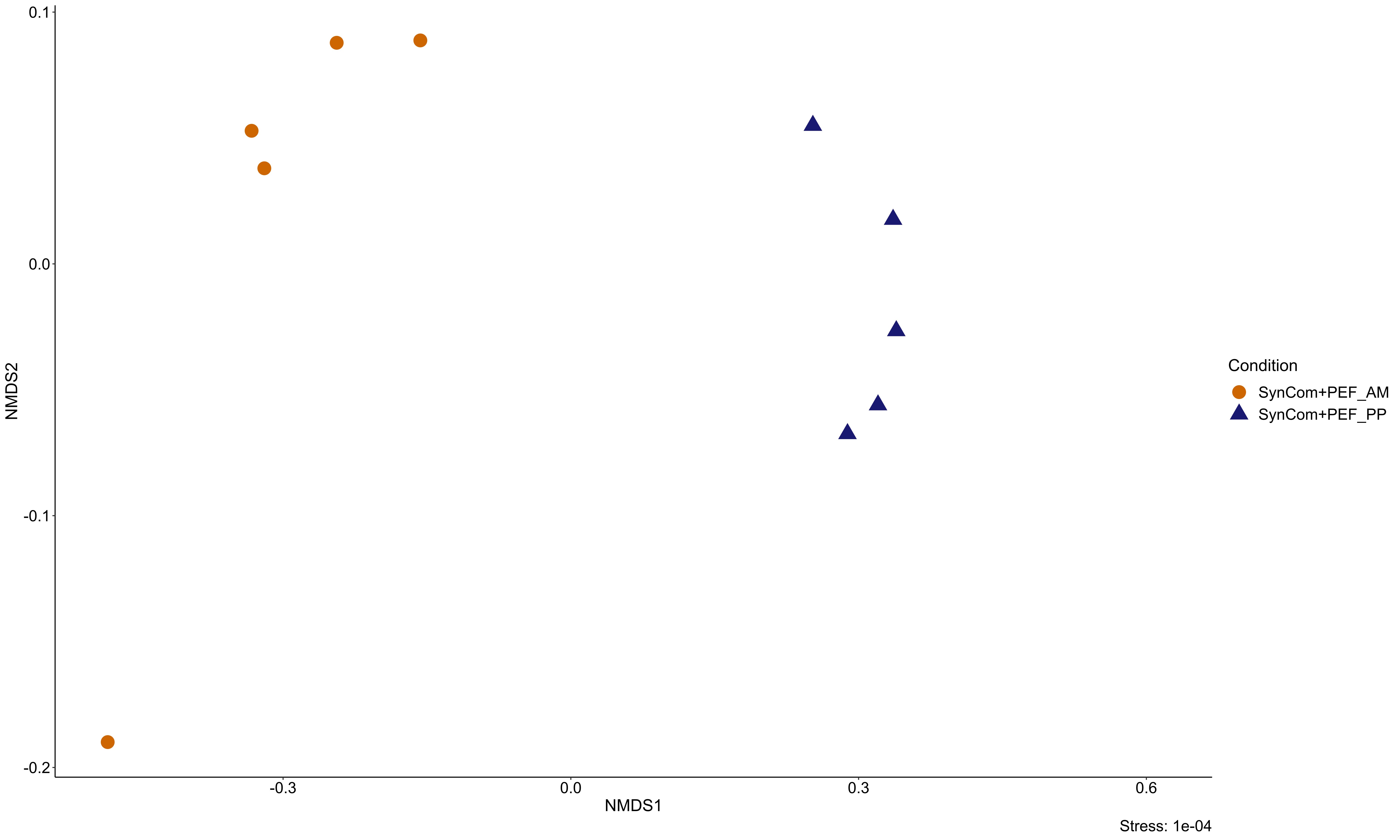
If PEF, irrespective of the producing algae, allowed the growth of copiotrophic strains, we found that exometabolites from the two different species led to distinct SynCom compositions. Hence, the PEF from A. minutum precluded the growth of some bacteria that were promoted when inoculated with P. parvum (i.e. Halomonas 09–027 and Marinovum). If competition for trophic resources potentially occurred between bacterial species in the SynCom, metabolic interactions likely participated in structuring the communities as well. Fu and collaborators (40) recently applied a different approach to study the role of phytoplankton exudates by growing natural bacterial communities in synthetic media mimicking phytoplankton-derived extracellular compounds. They found bacterial community assembly grown with several metabolites can be predicted based on the community composition grown with each of these metabolites. Our results are in agreement with these and support the role of algal exometabolites in favoring specific taxa (36, 38, 73). Cooperation may be another important biological process by which the growth of one partner depends on specific metabolic functions provided by the other members of the community, including nutrients or growth factors transfers (11), removal of toxic by-products (17) or degradation of complex carbon sources (5, 6). A well-known example of microbial metabolic interdependence is the synthesis of vitamins in auxotrophic communities, in which each member participates in the biosynthetic pathway by exchanging cofactor precursors that benefit the entire community (10). Such metabolic interactions may have occurred within SynComs, notably the one inoculated with PEF from P. parvum. While no growth of the bacterial isolates was observed, resource exploitation was more efficient in the presence of a complex and metabolically diverse bacterial community, which led to higher bacterial growth.
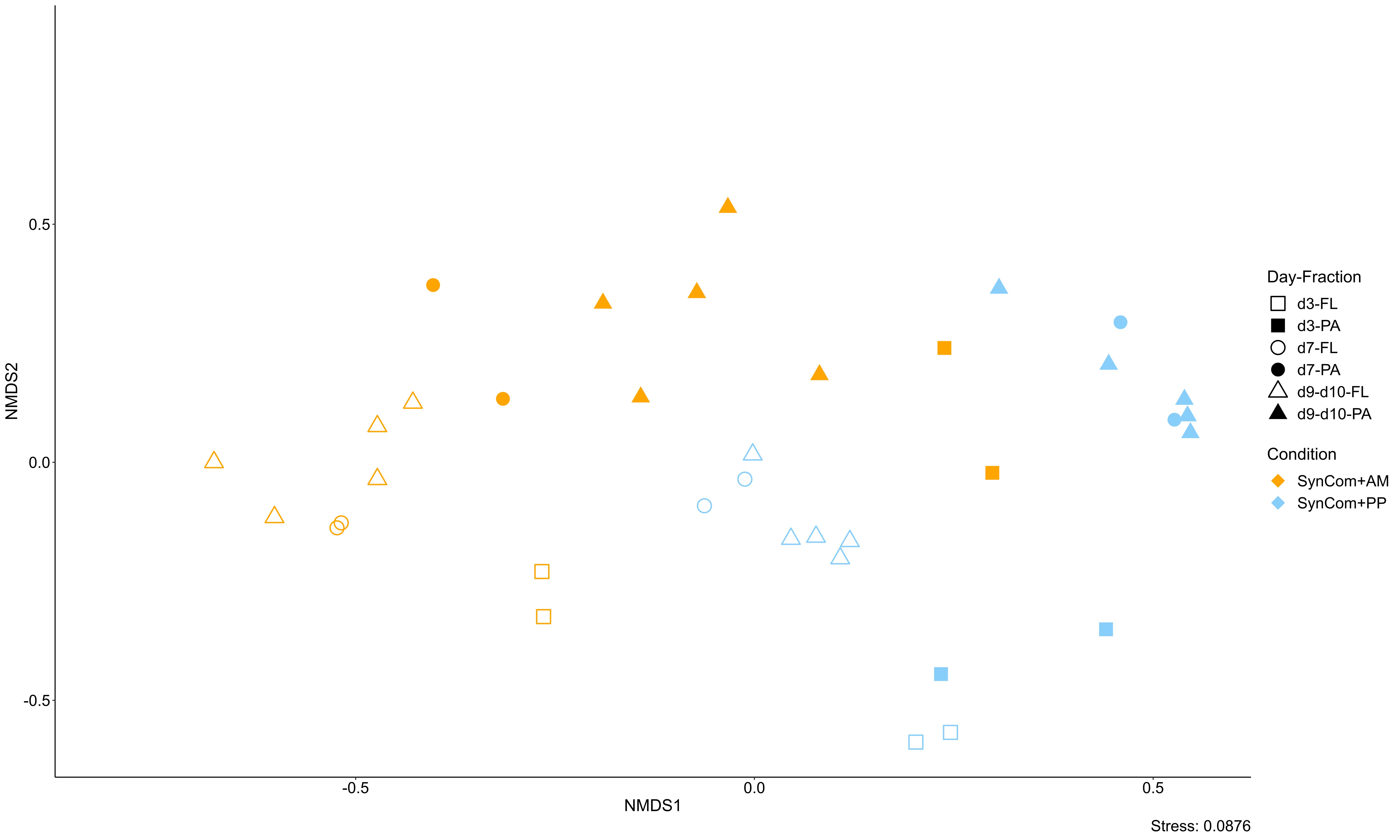
Moreover, as algae-bacteria interaction is two-way, our experimental design allowed us to discriminate between bacterial communities inoculated with PEF derived from the axenic algal host and the ones grown in contact with algal cells. Globally for each algal species, bacterial community analysis indicated high similarities between free-living (FL) and PEF populations and a higher specificity of particle-attached (PA) population, suggesting that the presence of algal cells also influenced the bacterial assembly. Thalassospira significantly enriched the FL fractions with higher relative abundance when associated with A. minutum. Bacteria of this genus are involved in the biodegradation of a variety of hydrocarbons (82–84). However, moving further into the interpretations would require deeper investigation into the chemical characterization of the A. minutum exometabolome. Conversely, Roseobacter and Marinovum were retrieved preferentially associated in PA fractions, suggesting an attached-lifestyle preference. Numerous studies have evidenced that particle-attached prokaryotes were phylogenetically and metabolically distinct from their free-living counterparts (23, 39, 85–87). Our observed differences may therefore be explained by the algal cells favoring the growth of bacteria that need attachment to grow and/or that bacteria may play an active role in modulating the production and release of algal metabolites in ways that optimize for bacterial growth. Here the SynCom was composed of bacteria isolated from phytoplankton cultures. These taxa, frequently found to be enriched in the phycosphere (8), may invest in mechanisms such as motility, chemotaxis and/or attachment to remain within the microenvironment (88). The enrichment of PA fractions by two members of the Roseobacter clade (Roseobacter and Marinovum) suggests that such molecular processes may have prevailed. This taxonomic lineage has been widely studied notably for the ability of some species to engage symbiotic interactions with phytoplankton cells (33, 89, 90). Moreover, recent studies evidenced that Roseobacters (Sulfitobacter and Phaeobacter) were able to switch from mobile to sessile lifestyles and that modulation of these molecular mechanisms were influenced by specialized exometabolites produced either by bacteria (N-acyl-homoserine lactone involved in quorum-sensing; (91)) or by diatoms (azelaic and rosmarinic acids; (42)). Exchange of infochemicals within the phycosphere is indeed an important driver of the relationship between microalgae and bacteria.
To conclude, we investigated the impact of exometabolites from different algal species on both bacterial isolates and microbial communities in presence and absence of the algal host. Using an artificial but ecologically relevant phycosphere microbial community, our work provides evidence that algal-specific extracellular compounds as well as the presence of algal cells were important drivers of the bacterial community composition. The mechanisms involved in this structuration are complex and include trophic competition, metabolic exchanges and chemical communication. This study paves the way for further investigations aiming to unravel the molecular processes involved in algae-bacteria and bacteria-bacteria interactions, and to understand the assembly, function and evolution of microbial communities. We encourage application of reductionist approaches to disentangle the inherent complexity of microbial interactions in the environment.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}