HSP60 expression was up-regulated in gout patients
It has been firmly established that HSP60 is secreted extracellularly and can enter the blood stream. Serum HSP60 levels are elevated in certain inflammatory diseases [11, 14]. In this study, to confirm HSP60 expression in patients with gout, HSP60 expression levels in serum and PBMCs were detected. Serum HSP60 levels were significantly higher in patients with AG than that in IG or HC (Fig. 1a). In PBMCs, HSP60 protein levels in gout patients were higher compared to HCs (Fig. 1b).
MSU crystals promoted HSP60 expression, translocation to mitochondria and secretion in RAW264.7 cells and THP-1-derived macrophage
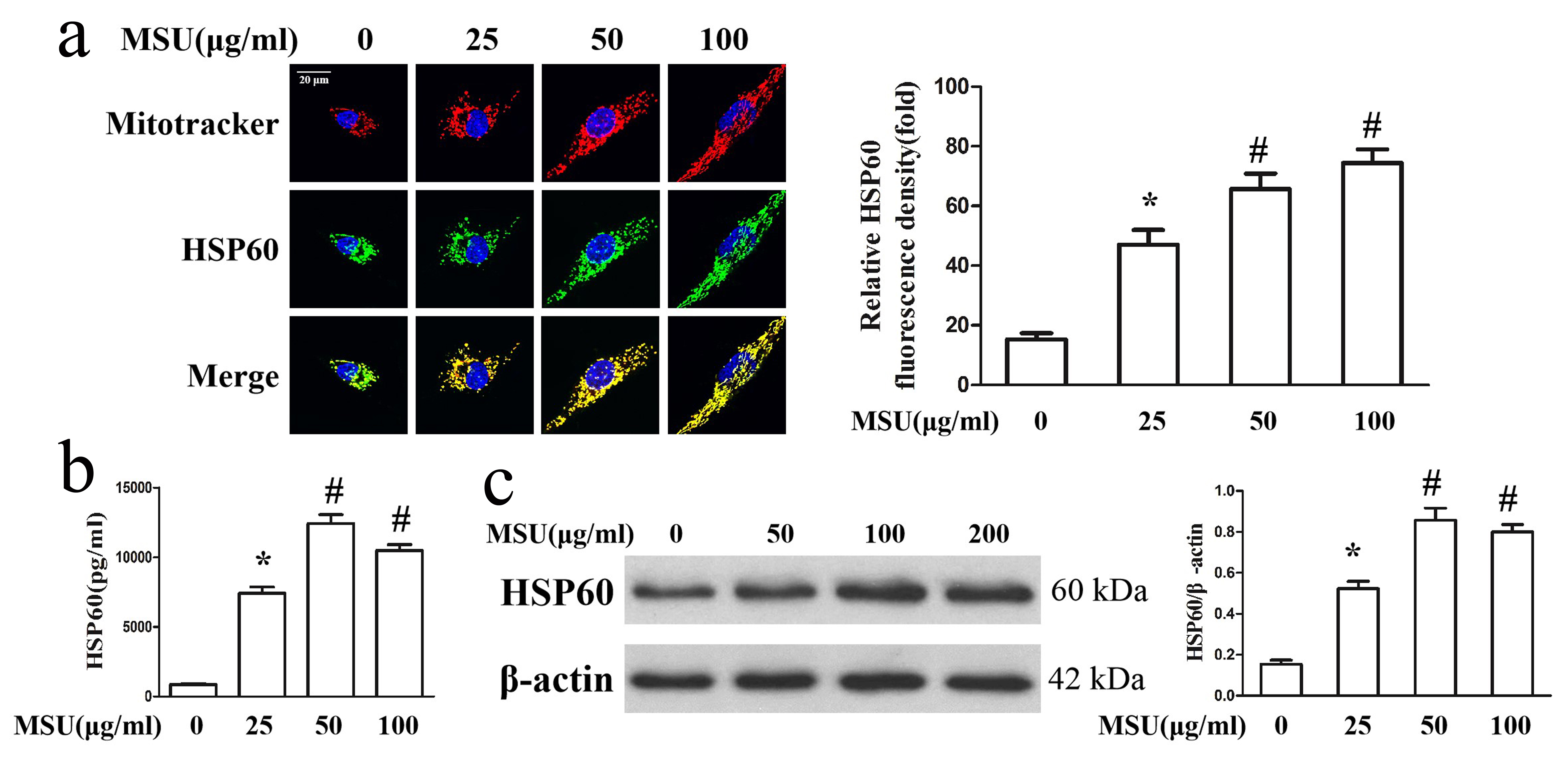
It has become clear that HSP60 is located not only in the mitochondria, but also in the cytoplasm that surrounding it. To investigate the effect of MSU crystal on the localization of HSP60 in the macrophage, RAW264.7 cells were exposed to varying dose of MSU crystals for 24 h, followed by an analysis of HSP60 distribution using double staining of MitoTracker and immunofluorescence. The double labeling of HSP60 and MitoTracker data revealed that HSP60 was predominantly located in the mitochondria and distributed through a small amount of cytoplasm in the macrophage; however, RAW264.7 cells exposed to MSU crystals, not only did HSP60 protein levels increase, but also it translocated to the mitochondria in almost its entirety (Fig. 1c). Next, we sought to examine the effect of MSU crystals on HSP60 secretion in RAW264.7 cells. Extracellular HSP60 levels were detected by ELISA in the culture supernatants, revealing an increased level of HSP60 secretion upon MSU crystals treatment (Fig. 1d). HSP60 expression levels were also assessed using western blot of protein lysates from RAW264.7 cells stimulated by different concentration of MSU crystals (Fig. 1e). MSU crystals dose dependently up-regulated the expression of HSP60 in RAW264.7 cells, and the expression of HSP60 peaked significantly in RAW264.7 cells treatment with 100μg/ml of MSU crystals. RAW264.7 cells were exposed to 100μg/ml of MSU crystals for all further assays. We further assessed the effect of different concentration of MSU crystals on the distribution, secretion and expression of HSP60 in the THP-1-derived macrophage. The results revealed that MSU crystals also had a great influence on the expression of HSP60, and HSP60 protein levels reached a peak in the THP-1-derived macrophage exposed to 50μg/ml of MSU crystals (Supplementary Fig. 1a, b and c). We chose to treat THP-1-derived macrophage with 50μg/ml of MSU crystals for all further experiments.
HSP60 regulates TLR4/MyD88/NF-κB signaling pathway in MSU crystal-stimulated inflammation
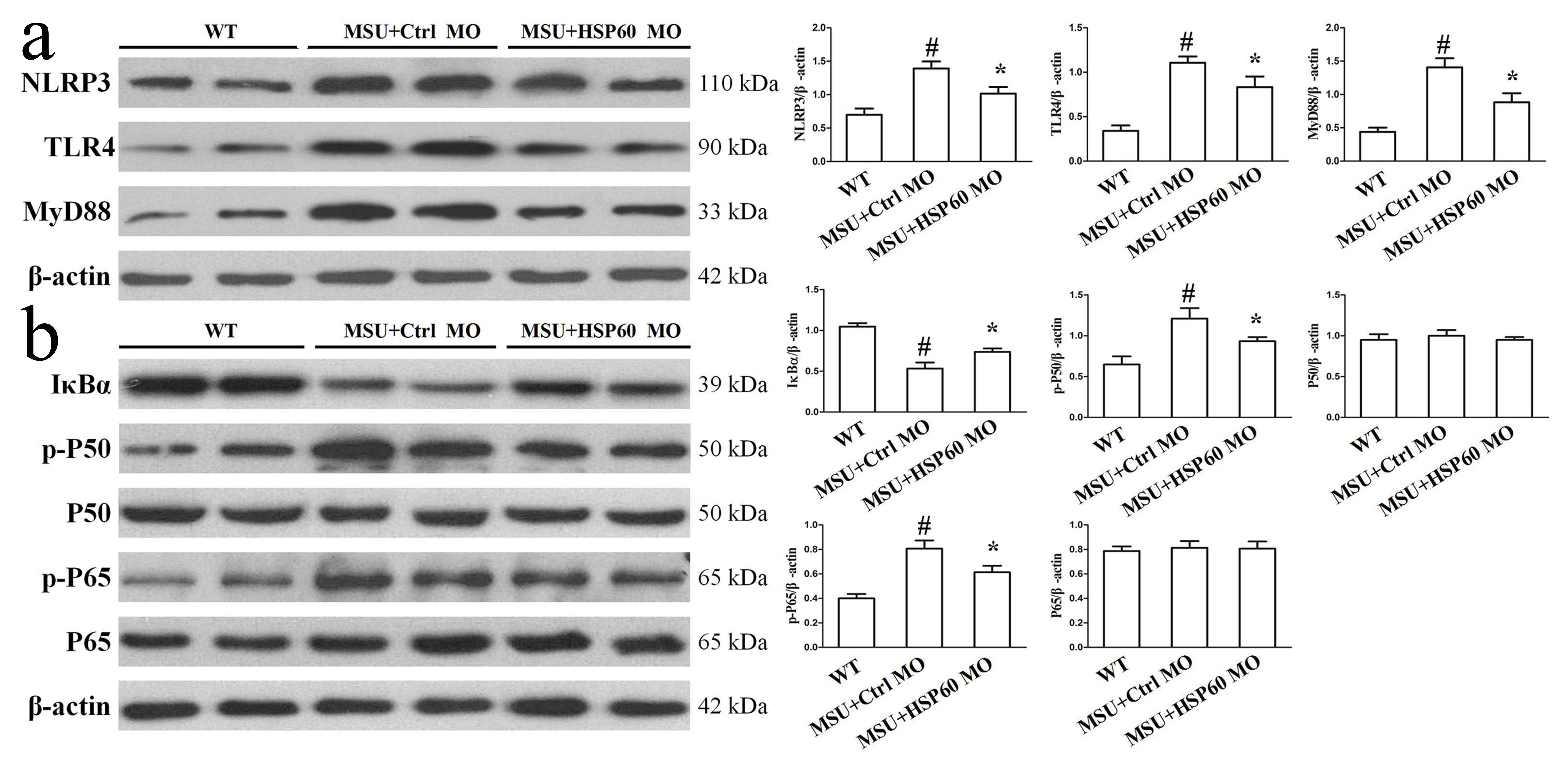
Previous studies have shown that HSP60 secretion into the extracellular medium activates TLR4 and induces an inflammation cascade [9, 15], and macrophages stimulated by MSU crystals also leads to the activation of the TLR4/MyD88 signaling pathway [16]. In our study, we set out to explore the effect of HSP60 on the TLR4/MyD88/NF-κB signaling pathway. We started by using specific siRNA to interfere with HSP60 expression in RAW264.7 cells for 48 h (Fig. 2a), followed by a 24 h MSU crystal treatment. Compared with control siRNA, the expression of TLR4 and its intracellular junction protein (MyD88) in RAW264.7 cells treatment with MSU crystals was inhibited due to down-regulation of HSP60 (Fig. 2b). The MyD88 signaling pathway is closely related to IκBα degradation and the phosphorylation of NF-κB in the MSU crystal-induced inflammation. As a result of HSP60 knockdown, the IκBα degradation and the phosphorylation of NF-κB P65 showed a reduction in MSU crystals exposed to RAW264.7 cells (Fig. 2c). For in vivo HSP60 knockdown, mice were injected with HSP60-Mo or control-Mo in the foot pad. Consistent with in vitro data, after HSP60 reduction by HSP60-MO, protein levels of TLR4, MyD88, phosphorylated NF-κB P65 and IκBα degradation decreased significantly in the foot pad tissue of MSU suspensions injection mice (Supplementary Fig. 2a and b).
Phosphorylation of NF-κB induces its nuclear localization, which is important for regulating inflammatory gene regulation. Therefore, we sought to evaluate the impact of both HSP60 knockdown and HSP60 overexpression on the nuclear localization of phosphorylated NF-κB P65 using immunofluorescence in RAW264.7 cells. Nuclear translocation of phosphorylated NF-κB P65 triggered by MSU crystals was attenuated by HSP60 knockdown (Fig. 2d). RAW264.7 cells were transfected with HSP60 vector or empty vector for 24 h and then treated with MSU crystals for 24 h. Western blot analysis was used to demonstrate HSP60 overexpression (Fig. 2e). HSP60 overexpression further accelerated the nuclear localization of phosphorylated NF-κB P65 induced by MSU crystals (Fig. 2f). These results imply that HSP60 plays a significant role in the MSU crystal-stimulated nuclear localization of the phosphorylated NF-κB subunit.
Both HSP60 knockdown and overexpression influence the production of downstream inflammatory cytokines of NF-κB induced by MSU crystals
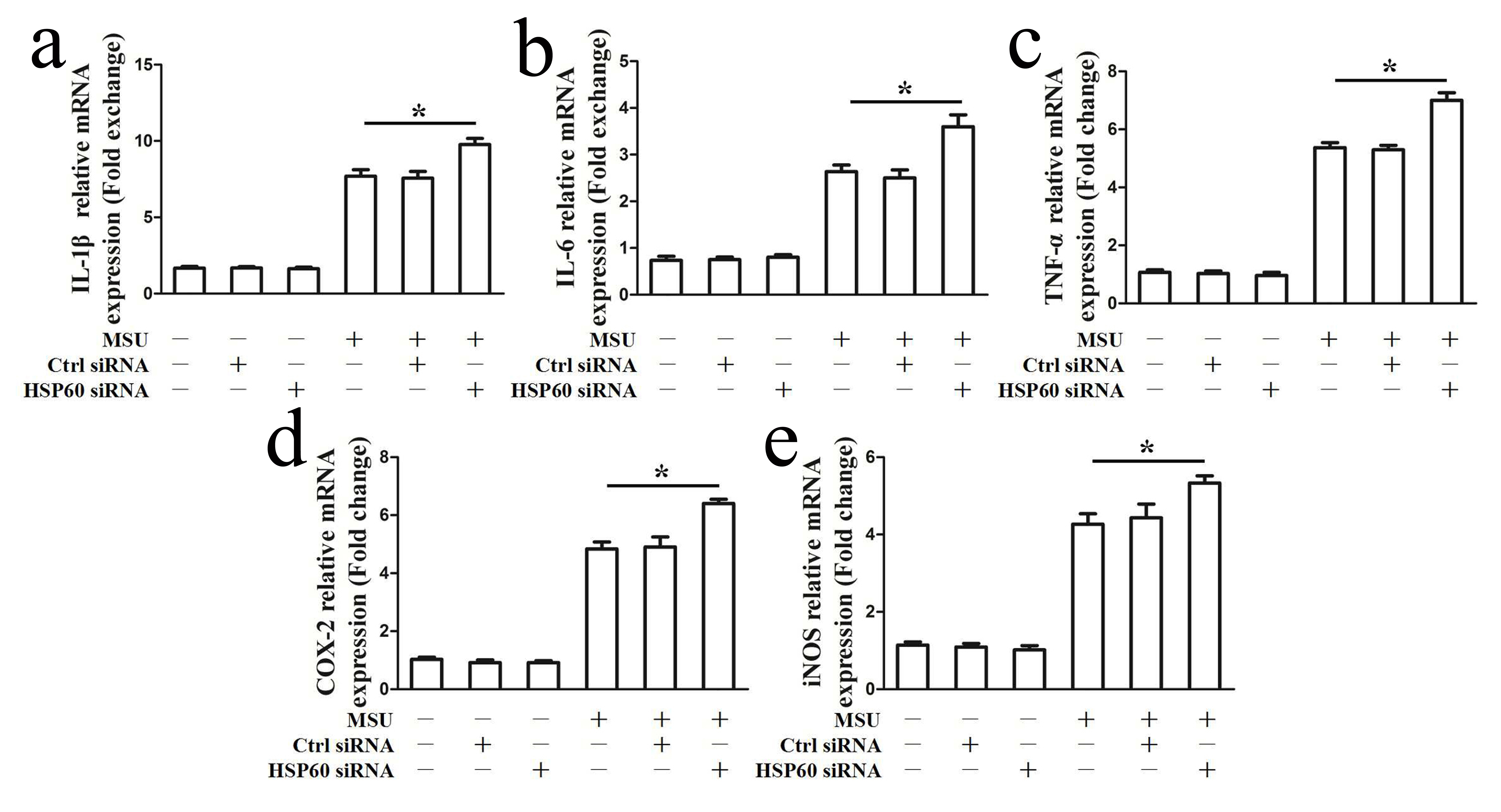
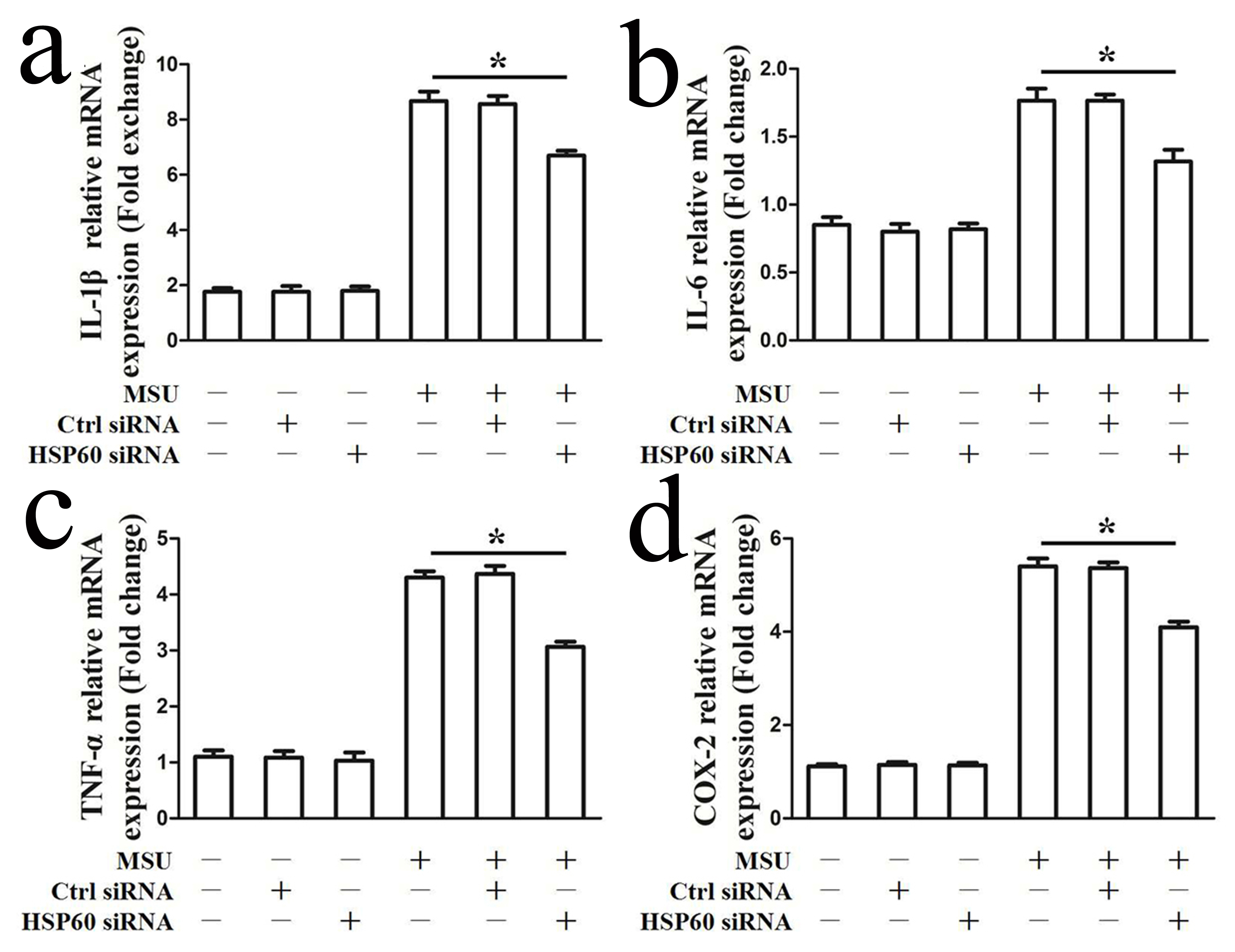
NF-κB activation impels the transcription of cytokines and prompts a complex inflammatory cascade response. As shown in Fig. 3a, reduction in endogenous HSP60 expression by specific siRNA in RAW264.7 cells blocked mRNA expression levels of MSU crystal-induced pro-inflammatory enzymes (COX-2, iNOS) and pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) (Fig. 3a). In accordance with the RT-PCR results, ELISA data indicated that lower IL-1β, TNF-α, IL-6 and PGE2 protein levels were found in the cell culture supernatants of HSP60 knockdown RAW264.7 cells in response to MSU crystals (Fig. 3b), and western blot analysis data also revealed that MSU crystal-induced iNOS and COX-2 protein levels were also suppressed in HSP60 knock-down RAW264.7 cells (Fig. 3c). To confirm this in human cells, we treated THP-1-derived HSP60 knockdown macrophages with MSU crystals. Consistently, HSP60 knockdown relieved the elevated mRNA expression of pro-inflammatory cytokines in response to MSU crystals (Supplementary Fig. 3). Inversely, we found that MSU crystals (100μg/ml) stimulated HSP60-overexpressing RAW264.7 cells significantly facilitating iNOS, COX-2, and pro-inflammatory cytokines (IL-1β, IL-6 and TNF-α) mRNA expression (Supplementary Fig. 4). All these data suggest that HSP60 has a positive effect on the production of MSU crystal-induced proinflammatory cytokines.
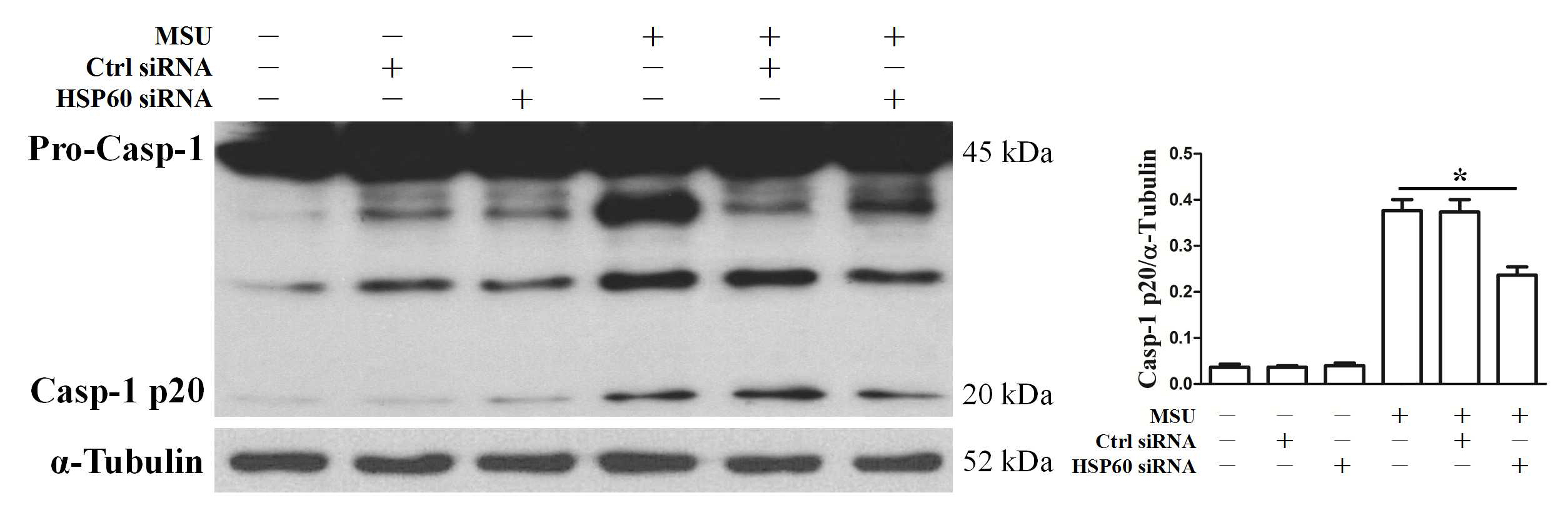
HSP60 regulates NLRP3 inflammasome activation triggered by MSU crystals
Previous studies have indicated that NLRP3 inflammasome activation is an important pathway by which MSU crystals lead to cellular inflammatory response, and nuclear localization of phosphorylated NF-κB promotes NLRP3 inflammasome activation by inducing transcription of the NLRP3 gene. Therefore, we investigated the role of HSP60 in NLRP3 expression induced by MSU crystals using RT-PCR and western blot. MSU crystal-induced NLRP3 expression both at mRNA and protein levels was decreased in vivo and in vitro because of HSP60 knockdown (Fig. 4a and b, Supplementary Fig. 2a). NLRP3 inflammasome activation contributes to the cleavage of pro-Caspase-1 to Caspase-1 and the release of IL-1β. The cleavage of pro-Caspase-1 to Caspase-1 and the production of IL-1β were alleviated because of HSP60 knockdown in the MSU crystals stimulated RAW264.7 cells and THP-1-derived macrophage (Fig. 4c, 4d, 4e, 4f and Supplementary Fig. 5). Oppositely, pro-Caspase-1 cleavage into Caspase-1 and the production of IL-1β were observed to be alleviated in HSP60 overexpression RAW264.7 cells stimulated by MSU crystals (Fig. 4g, 4h, 4i and 4j).
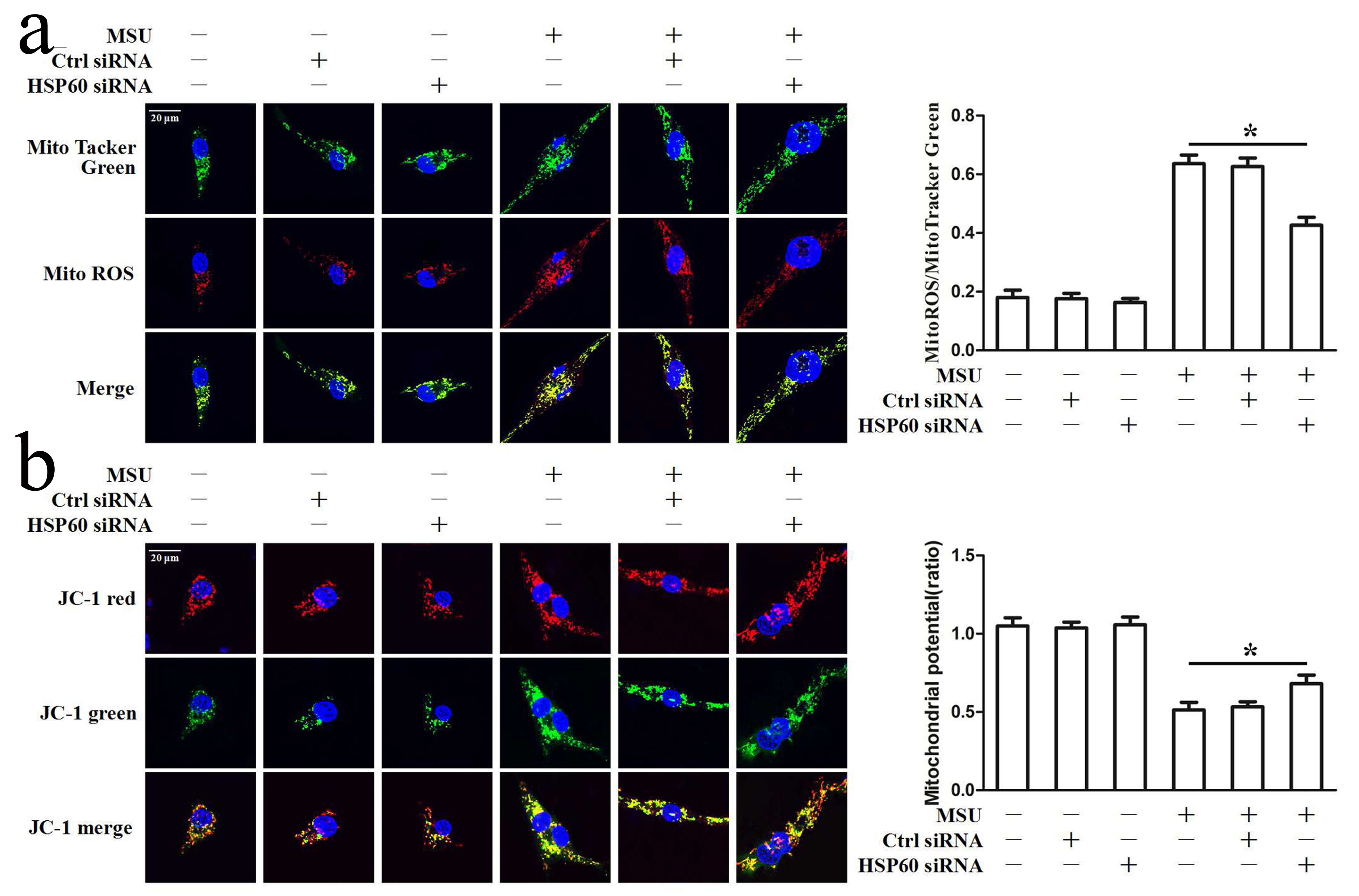
HSP60 aggravates mitochondrial dysfunction induced by MSU crystals
When mitochondria are damaged, HSP60 expression increased. HSP60 is considered an indicator of mitochondrial stress [17]. Mitochondrial dysfunction, which includes mitochondrial membrane potential collapse, overproduction of mitochondrial reactive oxygen species (mtROS) and release of mitochondrial DNA, is crucial for the activation of the NLRP3 inflammasome; therefore, we sought to explore the role of HSP60 in the mitochondrial damage induced by MSU crystals. MitoSOX Red is a mitochondrial superoxide indicator that is inserted into mitochondrial DNA during oxidation and produces red fluorescence. MitoTracker was used to determine the total mitochondrial content. Red to green fluorescence ratio reflects the mtROS level. HSP60 knockdown resulted in decreased MitoSOX Red fluorescence in MSU crystal-stimulated RAW264.7 cells and THP-1-derived macrophage, supporting lower levels of superoxide production (Fig. 5a and 5b, Supplementary Fig. 6a). Furthermore, the protein levels of mitochondrial matrix protein SOD2 and the activity of total superoxide dismutase (T-SOD) were displayed to be blocked in HSP60 knockdown RAW264.7 cells exposed to the MSU crystals (Fig. 5c and 5d), which further indicated that HSP60 has the function of regulating the production of mtROS induced by MSU crystals. We also found a reduction in mitochondrial DNA release to cytosol in HSP60 knockdown RAW264.7 cells in response to MSU crystals (Fig. 5e). In the study, MMP was measured by JC-1 probe. When the mitochondrial membrane potential is high, JC-1 gathered in the mitochondrial matrix and formed a red fluorescent polymer (J-aggregates); whereas the mitochondrial membrane potential is low, JC-1 cannot be concentrated in the mitochondrial matrix. In this case, JC-1 is a monomer and can produce green fluorescence. The ratio of red to green fluorescence is used to calculate mitochondrial membrane potential and reflects levels of polarized functional mitochondria. Because of HSP60 knockdown, mitochondrial membrane potential were restored in MSU crystal-induced RAW264.7 cells and THP-1-derived macrophages, supporting enhanced mitochondrial function (Fig. 5f and 5g, Supplementary Fig. 6b). On the other hand, we noticed that MMP collapse and mtROS generation triggered by MSU crystal-stimulated RAW264.7 cells were aggravated due to HSP60 overexpression (Fig. 5h, 5i, 5j, 5k).
HSP60 down-regulation alleviates the severity of MSU crystals induced mouse model of peritonitis and arthritis
We proceeded to study the role of HSP60 in the mouse model of gouty arthritis triggered by MSU crystals. After mice were respectively injected with HSP60-MO or ctrl-MO into the right foot pad tissue and right ankle joint for 1 h, MSU suspensions were respectively injected into the right ankle joint or right foot pad of mice to simulate the etiology of human gouty arthritis. A significant decrease in paw swelling index was noted in MSU+HSP60-MO treated mice compared to MSU+ctrl-MO treated mice (Fig. 6a). Both immunostaining and western blot data indicated that HSP60 expression levels were significantly lower in MSU+HSP60-MO treated mice compared to MSU+ctrl-MO treated mice (Fig. 6b and 6c). Histological analyses of the foot pad tissue section revealed that HSP60 knockdown prevented the infiltration of inflammatory cells into the foot pad tissue (Fig. 6d). We further confirmed the effect of down-regulated HSP60 expression on neutrophils, myeloperoxidases and macrophages distribution in the foot pad tissue section through immunostaining. As depicted in Fig. 6e, a decreased number of neutrophils and macrophages in the footpad tissue section were observed in the MSU+HSP6-MO treated mice. Concomitantly, the amount of MPO containing cells was also significantly lower in the MSU+HSP60-MO treated mice compared to MSU+ctrl-MO treated mice (Fig. 6e). We further assessed the protein levels of important pro-inflammatory enzymes (MPO, iNOS, and COX2) in the foot pad tissue extracts using western blot. The MPO, iNOS and COX-2 protein levels from the foot pad tissue extracts in HSP60-MO treated mice were found to be greatly lower (Fig. 6f). Compare to MSU+crtl-MO mice, the activity of MPO also revealed to be decreased in the MSU+HSP60-MO treated mice, reflecting lower levels of inflammation. HSP60-deficient mice had less ankle joint swelling compared to the ctrl vivo-morpholino treated group, which was consistent with the reduction in observed swelling of the foot pad (Fig. 6g). Mice were intraperitoneally injected with HSP60 for 1 h and then injected with MSU suspensions into the peritoneal cavity to establish a peritonitis model of gout. We found that total peritoneal cell numbers and IL-1β levels were lower in HSP60-MO treated mice (Fig. 6h and 6i). These results suggested that HSP60 knockdown relieved lymphocyte recruitment, alleviating MSU crystal-induced inflammation in vivo.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}