Currently, acupuncture has received widespread attention due to its remarkable efficacy in movement disorders, migraines, and chronic pain [24–26]. Modern medical treatment further developed acupuncture into EA therapy. Studies show that EA has a therapeutic effect on neurodegenerative diseases [27, 28]. Clinical studies have shown that EA could improve motor and non-motor dysfunction in PD patients [29]. However, the effect of EA stimulation on GV16, LR3, and ST36 acupoints to PD have not been reported. Therefore, we selected GV16, LR3, and ST36 as acupuncture points to verify the therapeutic effect of EA intervention on MPTP-induced PD mice in this study.
The occurrence of PD correlates with the patient’s body mass index. Studies reported that PD patients with decreased body mass index had more severe autonomic dysfunction and more rapid progression of motor deficits than those with normal, stable, or increased body mass index [30, 31]. It has also been demonstrated that gut microbiota disorders could promote PD progression. Increased intestinal bacteria, such as Lachnospira and Clostridia, contribute to weight loss and worsen PD symptoms [32]. In this study, we found that MPTP-induced PD mice showed a decreased body weight (Fig. 2a). Meanwhile, the body weight of PD mice increased after EA treatment. Besides, clinical studies have confirmed that WBC, RBC, HGB, MCHC, and LYM in the blood were down-regulated in PD patients [33]. In addition, lower Lym count is related to an increased risk of PD [34]. According to the blood routine test, we found the reduced counts of WBC, RBC, HGB, MCHC, and LYM in the MPTP group, which were elevated by EA (Fig. 2d–h). These data indicated a possible dysregulation of the neuro-immune-inflammatory network in MPTP-induced PD mice. In comparison, EA intervention might alleviate weight loss and blood routine dyscrasias in PD mice by modulating the neuro-immune-inflammatory network.
PD is triggered by the degeneration of dopaminergic neurons in the midbrain SN and striatum, and overexpression of α-syn in the midbrain impaired dopamine release and induced nigrostriatal neuronal cell death [35]. Therefore, the pathological accumulation of α-syn in the midbrain is considered a typical pathological feature of PD. In addition, PD occurs with decreased expression of TH, a key enzyme responsible for dopamine synthesis. At the same time, dopamine levels in the nigrostriatal pathway were reduced, leading to motor dysfunction [36]. In this study, we found that EA stimulation restored the motor function of PD mice (Fig. 1c − k). Besides, EA treatment increased TH expression in the midbrain and suppressed the protein levels of α-syn (Fig. 3e and 3f). Additionally, EA inhibited neuronal death caused by MPTP in the SN and striatum tissues (Fig. 2i and j). Therefore, it was suggested that EA intervention might improve motor dysfunction of PD mice by increasing midbrain TH levels, inhibiting disaggregation of α-syn, and protecting dopamine neurons.
Depression, one of the common comorbidities of PD, severely affects the quality of life in 40–50% of PD patients [37]. In Fig. 1c − k, the PD mice showed depression-like manifestations. BDNF, a member of the neurotrophic factor family, plays a crucial role in neuronal survival, synaptic plasticity, and depression [38]. Additionally, GDNF is closely connected to mood regulation and cognitive function. BDNF and GDNF levels elevated by amitriptyline could promote the survival of dopamine neurons and alleviate depression symptoms in PD [39]. In addition, researchers have also observed that the BDNF and GDNF expressions were reduced in the hippocampus and prefrontal cortex of PD animals [40]. Indeed, we observed that BDNF and GDNF were reduced in the midbrain of PD mice, an effect that was reversed by EA (Fig. 3d and 3e). Furthermore, EA treatment increased the viability and number of neuronal cells (Fig. 2i and j). Also, EA therapy improved depressive-like behaviors (Fig. 1c − k). Hence, it was implied that EA could improve the behavioral defects of PD mice by increasing the expressions of brain BDNF and GDNF and promoting neuronal survival.
Another significant complication of PD is intestinal dysfunction, which can start even more than twenty years before the motor symptoms. The activation of NLRP3 inflammasome induces the inflammatory response and disrupts intestinal tight junction proteins, leading to intestinal barrier disruption [41, 42]. In the present study, PD mice showed significant intestinal mucin disruption in the colon. Therefore, we examined NLRP3 inflammasome pathway-related factors in the colon tissues of mice. We found that the NLRP3 inflammasome was activated in the colon of PD mice, which was reversed by EA (Fig. 5a and 5b). Our data suggest that EA might alleviate intestinal inflammation and intestinal barrier damage by inhibiting the activation of colonic NLRP3 inflammasome. Nevertheless, the mechanism underlying the effect of EA initiated NLRP3 inflammasome inhibition remains to be discovered.
Dopamine contributes to gastrointestinal motility in the enteric nervous system. Thus, intestinal dopamine damage may lead to gastrointestinal dysfunction [43]. GDNF protects against MPTP-induced damage to the dopaminergic nervous system by inhibiting intestinal epithelial cell apoptosis, reducing intestinal permeability, inhibiting mucosal inflammation, and promoting intestinal barrier repair [44]. In addition, GDNF could promote the expression of Bcl-2, reduce the expression of Bax, and inhibit the activity of Caspase-3 [45]. In this study, we found that the transcription of GDNF was down-regulated in the colonic tissues of PD mice, which was increased by EA (Fig. 4d). Meanwhile, EA remarkably reduced the mRNA levels of Bax and Caspase-3 (Fig. 4f). Further, the levels of colonic glycoprotein and mucin were restored after EA treatment (Fig. 4a − c). These results suggest that EA could promote intestinal barrier repair by promoting GDNF expression and inhibiting apoptosis in PD mice.
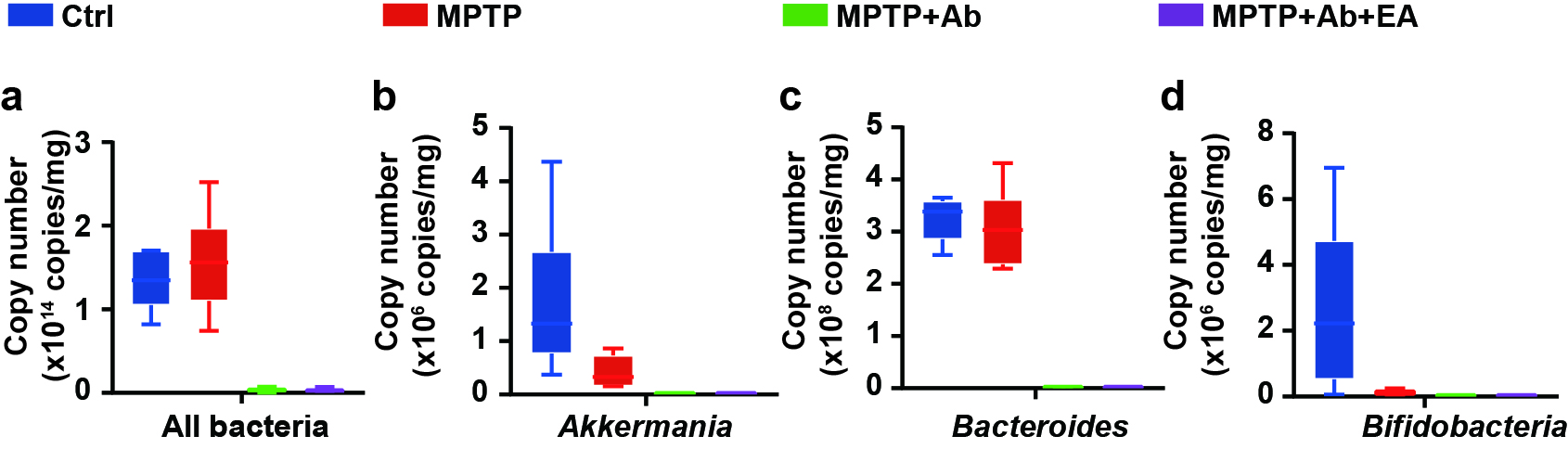
Intestinal inflammation in PD is associated with changes in intestinal permeability and microbiota composition[46]. Lai reported that the intraperitoneal injection of MPTP could induce dysbiosis in mice[47]. At the same time, probiotics alleviated the onset and progression of PD by regulating the structure and quantity of gut microbiota [48]. Han demonstrated that acupuncture stimulation could alter gut microbiota[49]. In this study, we found that the motor dysfunction and the up-regulation of NLRP3 and Cleaved-caspase-1 were improved by Ab-treatment compared to the MPTP group (Fig. 6 − 8). Similar results have been reported in other studies. For instance, Cui demonstrated that vancomycin pretreatment ameliorated motor dysfunction in MPTP-induced PD mice [50]. Another study found that the aggregation of alpha-syn and motor deficits was improved in mice with gut microbiota destruction by antibiotics[51]. Therefore, we believe that only a milder PD model can be established in antibiotic-treated mice (Fig. 6 and Fig. 7). The results show that EA failed to improve dopaminergic neurons, motor dysfunction, and intestinal inflammatory response in antibiotics-treated PD mice (Fig. 6–8). It points towards that EA may improve motor dysfunction and gut barrier integrity in PD mice by regulating gut microbiota.
{kind=link}