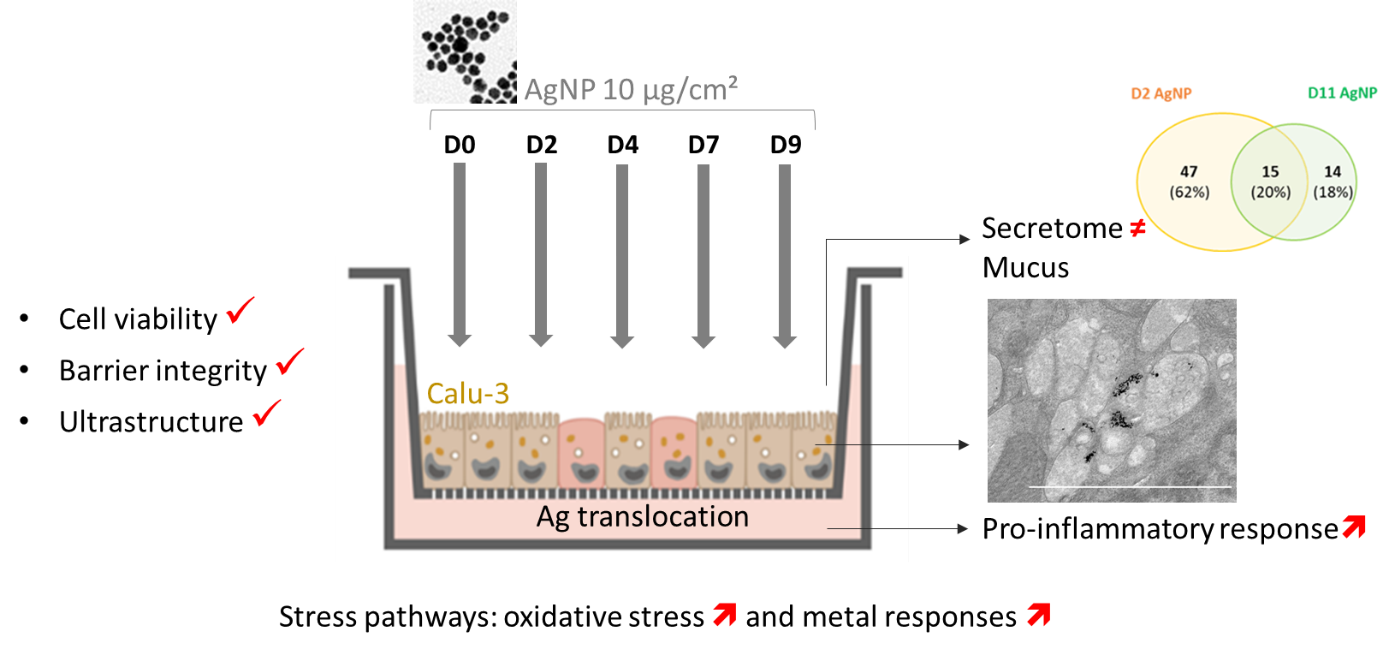
To explore the potential of the 3D Calu-3 model in assessing repeated toxicity, we set up an original exposure protocol (Fig. 1) and used AgNPs as model NMs. Experiments performed on 2D Calu-3 cell cultures showed an inhibitory concentration (IC50) of 20 µg/cm² for these nanoparticles, suggesting higher toxicity compared to other benchmark NMs tested within the European BIORIMA program. AgNPs are thus a relevant positive control for model validation in this study. 3D Calu-3 cultures were grown at ALI for 7 days before the first treatment, as we have previously shown that this lag time was necessary to obtain a well differentiated mature bronchial epithelium (Sanchez-Guzman et al. 2021). Cultures were exposed 5 times to 10 µg/cm² AgNPs every 2-3 days (D0, D2, D4, D7, D9) giving a cumulative nominal dose of 50 µg/cm². The effects were analysed 48 h after each exposure at D2, D4, D7, D9, and D11. Before each new treatment, the apical surface of the culture was rinsed with saline buffer (HBSSCa2+/Mg2+) to prevent AgNP accumulation and mimic mucociliary clearance occurring in vivo. Exposure was performed by deposition of 4 droplets of a low volume of AgNP suspension (20 µL/insert). This limited volume prevented detrimental effects of excessive apical liquid on the barrier integrity (Online Resource Fig. S1). We selected a set of noninvasive endpoints to monitor the cellular effects of repeated exposure of the bronchial epithelium to AgNPs without collecting or sacrificing cells (Fig. 1). This noninvasive strategy was completed with the analysis of gene expression from D2 to D11, and cell fixation at D11 for TEM observation.
2.1 Repeated exposure to AgNPs did not alter the permeability and viability of the Calu-3 barrier
Noninvasive assays were performed to evaluate the cytotoxicity and the alteration of the barrier integrity induced by AgNPs during repeated exposure. The Alamar Blue (AB) assay, which measures the cell metabolic activity, did not show any significant difference in cell viability as a function of time in control cultures (Fig. 2A). AgNPs did not induce a significant decrease in cell viability regardless of the number of treatments. A moderate but significant stimulation of metabolic activity was observed after treatment, except at D7 for which a 72 h recovery period was observed before analysis, as opposed to 48 h for the other time points. This increase could be due to an hormesis effect, that is a stimulation of the biological response following exposure to a low concentration of a chemical that is reducing the cell activity at higher concentrations. A similar hormesis effect has been described for A549 cell lines exposed to AgNPs, and was related to increased expression of heme oxygenase 1 (ho1) (Sthijns et al. 2017). The release of lactate dehydrogenase (LDH), which is an indicator of membrane damage, was measured in the basolateral culture medium and no significant increase of LDH was observed in exposed cells compared to controls (Fig. 2B). These results show that 5 repeated exposures to AgNPs at 10 µg/cm² did not induce cytotoxicity in the Calu-3 bronchial epithelium model.
Then, we controlled the barrier integrity by measuring the trans-epithelial electric resistance (TEER) (Fig. 2C) and Lucifer Yellow (LY) permeability (Fig. 2D). In controls, a decrease of the TEER was observed, but values remained above 300 W.cm² throughout the experiment, which corresponds to TEER values of a tight epithelium. In treated cells, a slight and significant increase of TEER was observed at D2, D4, and D9. The LY permeability assay confirmed that no damage or loss of barrier integrity was observed, as LY values remained < 2% in all conditions. The threshold values of 300 W.cm² and 2 % LY permeability were previously established for these culture conditions (Sanchez-Guzman et al. 2021). The same experiments were performed after 5 repeated exposures to 1 µg/cm² AgNPs. No cytotoxicity or alteration of the barrier integrity was observed (Online Resource Fig. S2).
2.2 AgNPs were internalized by epithelial cells without inducing ultrastructural alteration and Ag was translocated
Cultures were fixed at D2 and D11 to determine whether AgNPs were internalized and induced ultrastructural modifications. Observations of semi-thin sections stained with toluidine blue allowing an overview of the barrier structure did not show any major alteration of the epithelial barrier (Fig. 3A), as suggested by cell viability and membrane permeability assays. Surprisingly, some cells were observed on the basal side of the insert membrane, a phenomenon that we did not observe when we established the model (Sanchez-Guzman et al. 2021). Due to a manufacturing disruption of Corning inserts, we moved from Transwell® to Millicell® inserts for this study. Their analysis by Raman microscopy showed that both inserts share the same chemical composition, polyethylene terephthalate (PET). The porosity of the inserts was further characterized by Raman microscopy to analyze the pore morphology and to evaluate the percentage of single and fused pores in each insert (Online Resource Fig. S3A-C). A significant percentage of fused pores with a diameter > 3 µm pore size was observed for each insert type (40% and 38% fused pores for Transwell® and Millicell® inserts respectively). These larger pores may facilitate cell crossing. A similar pore density was measured for both inserts with 16,000 and 14,000 pores/mm² for Transwell® and Millicell® inserts respectively. Raman microscopy as well as observations of semi-thin sections revealed that the pore wall of Millicell® inserts was smoother, while it is more irregular and crenelated for Transwell® (Online Resource Fig. S3D-E), a feature that could favour cell crossing in Millicell® inserts. However, cell crossing seems limited after epithelial cell differentiation.
The ultrastructural observations of control and treated cells by TEM (Online Resource Fig. 3B, S4) did not show any visible effect after repeated exposure to AgNPs. Tight junctions, desmosomes, and microvilli were well visible in both conditions. Cells contained mucus granules and normal organelle ultrastructures were observed. In treated cells, AgNPs embedded in the mucus were observed extracellularly on the apical side of the culture, suggesting that some particles may be trapped in the mucus layer. Particles were also observed inside cells, often in vesicles, suggesting AgNP endocytosis (Fig. 3B). As several membranes were observed in these vesicles, they could be autophagosomes.
The translocation of AgNPs through the epithelial barrier was investigated by measuring total Ag concentration in the apical and basal medium after each exposure by ICP-MS (Online Resource Fig. S5). Because the analysis requires acidic dissolution of the sample, both Ag+ and AgNPs contribute to the total Ag concentration measured. Ag concentration in the apical medium increased between D2 and D9, suggesting that silver accumulated possibly due to decrease in cell uptake related to changes in mucus properties. The sum of the measured Ag concentration in the apical medium (163 mg/L) was lower than the nominal concentration (275 mg/L) corresponding to 5 repeated exposures at 10 µg/cm² (Online Resource Table S2), confirming partial consumption of AgNPs between treatments by cell uptake. Significant Ag concentrations were detected in the basal medium from D2 to D9, which demonstrates Ag translocation after each exposure. The highest Ag concentration was measured at D2 (51 µg/L) in the basal compartment, showing that silver (in ionic or particulate form) could translocate through the epithelium following the first treatment. The cumulated translocation was evaluated to 102.9 µg/L after 4 exposures, corresponding to 0.092 %, 0.031 %, 0.052 % and 0.012 % of the nominal silver concentration applied to the apical compartment after 1, 2, 3 and 4 exposures respectively and a cumulated percentage of 0.19 %. Even if the amount of silver inside cells could not be measured in this study, these data suggest that a fraction of AgNPs accumulate in the mucus layer where they could potentially be removed by mucociliary clearance especially as the concentration increase. Translocation results are also shown for 4 repeated exposures to 1 µg/cm² (Online Resource Fig. S5). A similar trend was observed with Ag translocation observed after the first exposure. A cumulated translocation of 19 µg/L (0.34 %) was measured. This decrease by a factor 5 compared to the experiment at high AgNP concentration is consistent with the measured Ag concentration in the apical compartment (Online Resource Table S2). These results clearly show that silver can cross the epithelial barrier. As the epithelium integrity is not compromised by AgNPs – tight junctions are preserved as shown by TEER and LY permeability – it suggests that translocation occurs by endocytosis of AgNPs followed by the release of Ag+ and/or AgNPs on the basal side of the cells within 48 h.
The translocation of Ag measured on Calu-3 bronchial epithelium model is similar to values reported in vitro after a single exposure to AgNPs of human Caco-2 intestinal epithelium (0.5 %) (Bouwmeester et al. 2011) and of human co-culture model of lung Calu-3 epithelial and EA.hy926 endothelial cells (1 %) (Zhang et al. 2019). The penetration of AgNPs in the human nasal epithelium was also reported ex vivo, as well as Ag translocation through nasal epithelium model in vitro (Falconer et al. 2018), suggesting that a risk of silver translocation exists in case of exposure to AgNPs by inhalation at the different airway levels.
2.3 AgNPs induced a pro-inflammatory response without mucus hypersecretion
In order to evaluate whether AgNPs induced a proinflammatory response, IL-6 and IL-8 proinflammatory cytokines were measured in the apical and basolateral media after each exposure. For an accurate interpretation of these data, the evolution of the constitutive release of these cytokines was first analyzed during the experimental time (12 days from D0 to D11) (Fig. 4). IL-8 was released in greater amount than IL-6 (15,000 pg and 600 pg respectively at D0) (Fig. 4A). Both apical and basal releases were observed for IL-6 and IL-8 (Fig. 4B-C). The release of IL-6 was polarized towards the apical side (70% of total secretion), whereas the release of IL-8 was equivalent in both orientations (Fig. 4B-C). The concentrations of IL-6 and IL-8 tended to increase with time in the apical medium in control cells, but they remained stable, except at D2, in the basal medium (Online Resource Fig. S6).
Then, we checked whether cytokines adsorbed on AgNPs, which could interfere with the measurement of their concentration in treated cultures (Online Resource Fig. S7). The results showed that AgNPs interfered with the ELISA assay. Therefore, we analyzed IL-6 and IL-8 release by treated cells only in the basal medium. Repeated exposure to 10 µg/cm² AgNPs induced a significant increase of IL-8 release in the basal medium regardless of the number of exposure, whereas no increase was observed following repeated exposure to 1 µg/cm² (Fig. 5A). Neither adaptation nor exacerbation of the pro-inflammatory response was observed after repeated exposure compared to single exposure to AgNPs. Similar results were observed for IL-6 release in the basal medium to a lesser extent (Fig. 5B). The RT-qPCR results showed no increase in mRNA expression of these two cytokines (Fig. 5C-D).
Finally, mucus secretion was analyzed to determine whether AgNPs induced mucus hypersecretion. This potential NM effect was indirectly assessed using the ELLA assay that quantifies glycoprotein content (as mucins are the most abundant glycoproteins in the secretome). The constitutive release of glycoproteins in the apical secretome remained constant during 12 days (Fig. 6). In treated cultures, a significant decrease of glycoprotein concentration was observed after 3 and 4 exposures (17% and 14 % decrease respectively). In order to determine whether this decrease was consistent with a reduction of mucin mRNA expression, MUC5AC expression that corresponds to mucin5ac, one of the main mucins, was analysed. No significant change of MUC5AC expression was observed regardless of the number of treatments (Online Resource Fig. S8). These results suggest that either repeated exposure to AgNPs decreased the mRNA expression of other mucins, or decreased glycoprotein secretions without altering their mRNA expression.
2.4 AgNPs induced antioxidant and metal stress responses of the epithelial cells
To determine if oxidative stress is a toxicity pathway of AgNPs, we first evaluated the intrinsic oxidative potential of the particles by measuring the depletion in antioxidant molecules, ascorbic acid (AA), uric acid (UA), and gutathione (GSH), of a simplified lung lining fluid (Crobeddu et al. 2017). Little or no depletion of AA, UA, GSH (< 10%) was observed after 4 h incubation with AgNPs at concentrations up to 10 µg/mL at 37°C (Online Resource Fig. S9). By contrast, high depletion of AA and GSH (100%) was observed for copper nanoparticles in the same conditions (Online Resource Fig. S9). These results suggest that AgNPs used in this study have limited short term acellular oxidant effects compared to other metallic nanoparticles. To evaluate the cellular oxidative stress response, we measured after each exposure, the expression of heme oxygenase 1 (HO-1), an antioxidant enzyme expressed by Calu-3 cells and regulated by the Nrf2 (Nuclear factor erythroid 2-related factor 2) transcription factor, (Fig. 7A). A significant induction of HO-1 was observed 48 h after each treatment, while no induction was observed 72 h after the 3rd treatment, suggesting a transient induction of HO-1 that is restored 72 h after exposure. HO-1 transient expression, which is typical of adaptive oxidative stress response, was previously observed in bronchial epithelial cells exposed to ambient particles (Val et al. 2011).
In the case of metal stress, the adaptive cell response involves the activation of the metal regulatory transcription factor 1 (MTF1) that controls the expression of metallothionein 1X (MT1X) and 2A (MT2A) ) which will bind the metals that led to the activation of the transcription factor. MT1X and MT2A expressions were significantly induced in treated cells both 48 and 72 h post exposure (Fig. 7B-C). The increase of metallothionein expressions following consecutive exposures with a 48 h interval (D4 > D2, D11 > D9 > D7) and the lower level of induction observed after 72 h (D7 < D4) suggests that the induction of metallothioneins in epithelial cells leads to the successful sequestration of Ag+ ions when the period between two successive treatments is long enough. As the induction of MT1X and MT2A was still significant at D7 compared to controls, we conclude that 72 h were not sufficient for cells to completely deal with the metal stress. Because the activation of MTF1 transcription factors requires binding of Ag+ to MTF1 (Luther et al. 2012), this effect could be due to slow dissolution kinetics of AgNPs after endocytosis.
The bronchial epithelium was exposed to silver nitrate (AgNO3) to compare the effects of exposure to silver ions and AgNPs. 2 repeated exposures were performed at D0 and D2 following the same protocol with an equivalent Ag concentration of 10 µg/cm². No effects of repeated exposure to AgNO3 on Calu-3 cell viability and barrier integrity were observed (Online Resource Fig. S10A-C). We did not observe any induction of the expression of IL-6, IL-8, and MUC5AC (Online Resource Fig. S10D-F). However, a significant induction of HO-1, MT1X, and MT2A expressions was observed (Online Resource Fig. S10G-I). A stronger induction of metallothionein genes was observed following a single exposure to AgNO3 compared to AgNPs. A lower induction was observed after the second exposure to AgNO3 in contrary to AgNPs, which suggests that AgNP dissolution is involved in the delayed metal stress response observed in epithelial cells following multiple exposure. We have shown previously by SAXS analysis that AgNP dissolution in MEM is very limited after incubation for 18 days at 37°C (Martin et al. 2022). Therefore, it is likely that AgNP dissolution occurs after endocytosis in the phagolysosomes observed by TEM, where dissolution would be favoured by low pH. The dissolution of AgNPs in lysosomes after endocytosis has been reported for different cell types in vitro. However, the dissolution kinetics of AgNPs depends on the particle shape, surface area, and capping or stabilizing agent, which in turn modulates the antioxidant and metal stress response of the cells (Graf et al. 2018) (Smith et al. 2018) (Gonzalez et al. 2015).
2.5 Single and repeated exposure to AgNPs induced different changes in secretome composition
In addition to cytokines and mucins, Calu-3 cells secrete numerous proteins on their apical side, which contribute to the immune response and cell-to-cell communication of the bronchial epithelium (Sanchez-Guzman et al. 2021) (Gupta et al. 2019). We analyzed the composition of the apical secretome of Calu-3 cells by quantitative proteomics to determine whether single and repeated exposure to AgNPs modulate the secretome composition, and whether these secreted proteins are involved in the cellular defense. The secretome composition was analyzed at D2 after a single exposure (‘D2 AgNP’) and at D11 after 5 repeated exposures (‘D11 AgNP’), and compared to the secretomes of control cells at the same time points (‘D2 Control’, ‘D11 Control’) (Fig. 8). More than 500 extracellular proteins were identified in the apical secretome of Calu-3 cells, which is comparable to our previous results on the long-term evolution of the bronchial epithelium secretome (Sanchez-Guzman et al. 2021). The Principal Component Analysis (PCA) performed on the 4 conditions showed two separate clusters for D2 AgNP and D11 AgNP respectively, suggesting differences in the secretome after single or repeated exposure to AgNPs (Fig. 8A). The comparison with the secretome of control cells is shown as a Venn diagram (Fig. 8B). The secretomes of D2 Control and D2 AgNP share 432 common extracellular proteins, while 62 proteins are specifically secreted by cells exposed to AgNPs (12 %), and 9 proteins are secreted by control cells only (2 %). The secretomes of D11 Control and D11 AgNP share 463 common extracellular proteins, while 29 proteins are specifically secreted by cells exposed to AgNPs (6 %), and 12 proteins are secreted by control cells only (2 %). We conclude that AgNPs induced secretion of specific proteins by epithelial cells at D2 and D11. The evolution of the secretome composition with time for control cells shows that 87 % of the secreted proteins are common at D2 and D11 (Online Resource Fig. S11A). Interestingly, when comparing proteins that are secreted by exposed cells only, we can see that 47 proteins are secreted at D2 AgNP only (62 %), 14 proteins are secreted at D11 AgNP only (18 %), and 15 proteins are common to D2 AgNP and D11 AgNP (20 %) (Fig. 8C). This result demonstrates that single and repeated exposure to AgNPs leads to different responses in terms of protein secretion by epithelial cells.
To further investigate this effect, the reactome enrichment pathways of the secretomes were analyzed (Fig. 8D). Significant pathways (p > 0.5) identified in D11 AgNP were not expressed in D2 AgNP, namely apoptosis, programmed cell death, and suppression of autophagy. As no loss of cell viability was observed during AgNP exposure, it suggests that these processes could be related to the cellular turnover of the cellular layer by anoikis, which has already been observed with this model (Sanchez-Guzman et al. 2021). Secreted proteins involved in neutrophil degranulation were strongly expressed at D2, but not at D11, suggesting a stronger immune response after a single exposure.
The composition of the 20 most abundant proteins in the secretome of control and exposed cells was compared using a heat map to highlight differences in secreted protein abundancy and identify depleted or enriched proteins in the secretome after exposure to AgNPs (Online Resource Fig. S11B). α1-antichymotrypsin (serpina3) and galectin-3 binding protein (lgals3bp) proteins were enriched in the secretome of D2 AgNPs, suggesting enhanced enzymatic activity and immune response of the epithelium after exposure to AgNPs, while mucin 5AC (muc5ac) was depleted compared to control cells, in line with our observation on glycoprotein secretion. Polymeric immunoglobulin receptor (pigr) and immunoglobulin G (fcgbp) were both enriched in the secretome of D11 AgNPs, suggesting that epithelial immune response was also triggered after repeated exposure to AgNPs. The absence of pigr enrichment at D2 AgNP after a single exposure could be explained by the long-term evolution of the epithelium in vitro, as we observed before that this protein becomes more abundant in the mature secretome of Calu-3 cells (Sanchez-Guzman et al. 2021). By contrast, α1-antitrypsin (serpina1) was depleted in the secretome of D11 AgNP compared to D11 Control and D2 AgNP, suggesting that the enzymatic balance between proteases and protease inhibitors in the secretome may change after repeated exposure to AgNPs. The analysis of the secretome revealed changes in the secreted protein profiles associated with the epithelial immune response after multiple exposure to AgNPs that were unseen after a single exposure to the toxic compound of interest.
2.6 Advantages and limits of Calu-3 models for chronic toxicity studies
In this study, we used Calu-3 cell line to generate a human bronchial epithelial barrier. Calu-3 cells are commercially available, which is an advantage compared to NHBE (normal human bronchial epithelial cells), especially for screening different compounds or conditions. Primary cells from donors are available at higher cost and present high variability between individuals, which requires repeating the experiments with epithelia from different donors. In the context of the development of Integrated Approaches to Testing and Assessment (IATA), a low cost model present advantages when used in the first tier of tests to identify toxicity mechanisms, which can then be confirmed using NHBE models. The Calu-3 model was also used by Braakhuis et al to study particle toxicity following aerosol exposure using a continuous airflow and an automated exposure system (Braakhuis et al. 2020). This device reproduces realistic exposure conditions, but it requires expensive equipment. The aerosolization method is not applicable to all inhaled contaminants due to the heating process which is not compatible with some organic material or plastic particls. We demonstrate here that the pseudo-ALI method we used for exposure allowed 5 repeated exposures without any damage to the epithelial barrier. One experiment was performed with 12 repeated exposures during 28 days to reproduce in vitro the sub-acute toxicity experiments performed during 28 days in vivo (data not shown). Our results also highlight the importance of the insert membrane choice to limit cell crossing. This simple and low-cost model compared to primary cell cultures can be easily used by laboratories to address the issue of chronic toxicity.
Using this model, we assessed the toxicity of AgNPs on the bronchial epithelium following repeated exposure during 12 days. We observed a pro-inflammatory, antioxidant and metal stress response in vitro that is consistent with in vivo data. Neutrophil infiltration of the bronchoalveolar lavage of mice (Stebounova et al. 2011) or rats (Braakhuis et al. 2014) was observed after AgNP inhalation for 2 weeks and 4 days respectively. Moderate inflammation with lung damage and transient neutrophil infiltration was already shown after a single intratracheal instillation of mice to AgNPs (Kaewamatawong et al. 2014). It was associated to oxidative stress and metal stress with the induction of SOD and metallothioneins in alveolar cells, in a transient manner which is consistent with the results obtained in this study. AgNPs internalization observed here was also reported in vivo in lung epithelial cells (Braakhuis et al. 2014) (Sung et al. 2009).
Our model allowed us to observe in vitro transient effects and epithelial cell responses that are increased with multiple exposures. This model is thus well adapted to study the adaptive response of the epithelium to repeated and long-term exposure to pollutants. As in vitro responses after repeated exposure to AgNP are consistent with in vivo observations, it demonstrates the suitability and reliability of this model for toxicology studies.
{kind=link}