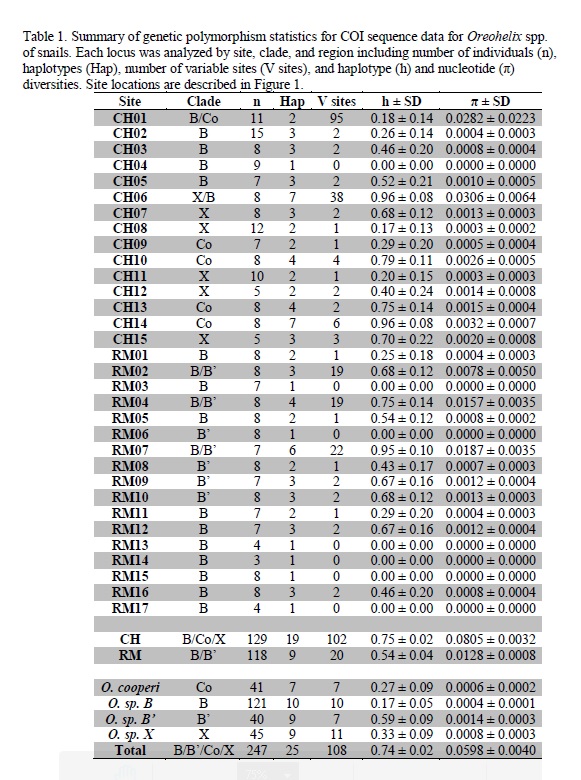
We obtained Oreohelid snails from 58 sites in two areas of southern Alberta and Saskatchewan. Two closely-related mitochondrial DNA clades were found in the Rocky Mountains, and three clades were found in Cypress Hills. Of the four COI clades identified, only Oreohelix cooperi was morphologically distinguishable [13, 16, 17]. The larger snails have been identified as belonging to the species complex Oreohelix subrudis, and are widespread throughout the Rocky Mountains and sky islands in Montana and Wyoming [10]. Two of the lineages sampled in Canada had highly restricted ranges: Clades X and B’. Clade X has only been found in Cypress Hills and is most closely related to Clade E in Wyoming described by [10] indicating that Clade X is either a glacial relict in Cypress Hills or colonized Cypress Hills from the periglacial zone immediately south of Cypress Hills and the Laurentide ice sheet. Clade B’ is restricted to the northern end of the sampled range in the Rocky Mountains and is closely related to Clade B. This proximity suggests that Clade B’ recently diverged from Clade B in the Rocky Mountains. The presence of isolated lineages restricted to a small range as well as widespread lineages is concordant with findings from [10] on Oreohelid snails collected further to the south.
High levels of genetic divergence at the mitochondrial locus and cryptic speciation are commonly reported in studies of terrestrial snails [18]. [19] attributes the high divergence to one of four possibilities: (i) relatively rapid mitochondrial divergence, (ii) prolonged isolation followed by secondary contact, (iii) selection pressure to maintain multiple mitochondrial clades, or (iv) colonization patterns leading to the co-occurrence presence of many divergent mitochondrial clades. The COI patterns observed in the snails sampled are best described by a combination of (ii) and (iv). Secondary contact following prolonged isolation (ii) best describes the presence of the three divergent mitochondrial lineages in Cypress Hills, particularly Clades B and X where nuclear ITS data show limited mixing. One possibility is that each of the clades of the O. subrudis species complex evolved in isolation on a sky island. Temperature and precipitation changes characterizing the Pleistocene glaciations allowed some of these clades to come into secondary contact. During the temperature and precipitation fluctuations in the early Holocene [20, 21], Cypress Hills and other sky islands such as the Black Hills or Bighorn Hills remained as relatively stable habitat for Oreohelids [22]. This effect may be especially pronounced in a few areas, such as Cypress Hills and the Bighorn Mountains (WY1, WY2, and WY3 from [10]). Secondary contact following prolonged isolation has been found in the dusky Arion slug (Arion subfuscus; [23]). [23] reported highly divergent mitochondrial sequence data with low nuclear ITS1 divergence, which matches the pattern we found in our Oreohelid snails. [23] suggests that hybridization of nuclear loci occurred in the slugs, leading to the maintenance of multiple divergent mitochondrial lineages.
Alternatively, colonization patterns leading to many divergent mitochondrial clades (iv) could account for the overall patterns of distribution of O. cooperi and O. subrudis in North America, including Cypress Hills. Under this model, an area is colonized either actively through a habitat corridor or through passive long-range dispersal [24-26]. The new population quickly expands in size, thereby reducing the likelihood of further invasion. Colonization patterns favouring multiple divergent mitochondrial clades were found to be the best explanation for the distribution of mitotypes in grove snails (Cepea nemoralis) across northwestern Europe [19] due to their high population sizes and distances between patches of suitable habitat. These authors argued that populations of grove snails are effectively arranged like stepping stones, and dispersal events are rare.
ITS2 and cytonuclear discordance
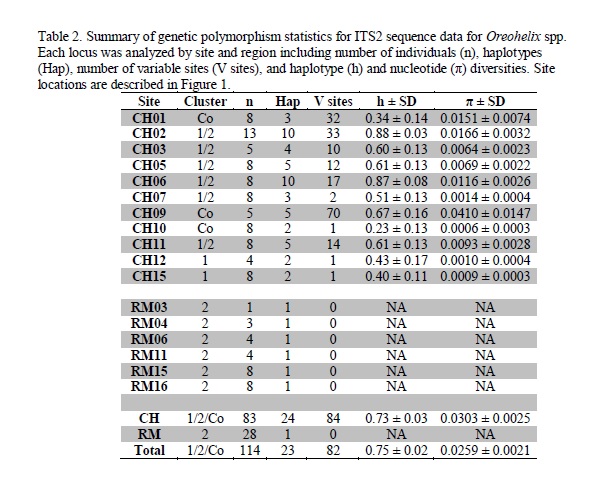
ITS2 data revealed three genetic clusters; one in the Rocky Mountains, and three in Cypress Hills. One cluster was exclusive to O. cooperi. These results confirm those from our earlier study showing O. cooperi is genetically distinct from other sympatric Oreohelids and it is endemic in Canada to the Cypress Hills [16]. Snails in Cluster 1 were typical of O. sp. X and cluster 2 was typical of O. sp. B, but the two divergent mitochondrial clades showed overlap at the ITS2 locus, particularly surrounding the contact zone in the Cypress Hills. There are two possibilities why Clades B and X exhibit mitochondrial and nuclear discordance. The first possibility is incomplete lineage sorting, and the second is introgression and hybridization [27]. While these are not mutually exclusive, hybridization and introgression better explain the ITS2 pattern observed in Cypress Hills. If incomplete lineage sorting was occurring, there would be no geographic pattern to the nuclear allele distribution in Cypress Hills [28]. However, there is a clear geographic pattern to both the mitochondrial COI and the nuclear ITS2. While both alleles are common in a small area surrounding the contact zone near Elkwater Lake, to the east of the contact zone near Highway 41 alleles belonging to Clade 1 become much more common.
Glacial history
Oreohelids have a fossil record in North America dating back to the Cretaceous period [29], but the current distribution of Oreohelids in southern Alberta is a consequence of the Quaternary glacial history of the region. During the LGM, Cypress Hills was inaccessible to terrestrial snails and surrounded by ice. As the ice sheets receded, the southern slopes were the first to become ice free and allow access to the hills, while the northern slopes retained ice for much longer [30]. The earliest indication of vegetation in Cypress Hills is approximately 9000 years before present [22] based on pollen data from Harris Lake (49.6663 ºN 109.9044 ºW) on the northern side of Cypress Hills. Literature is sparse regarding the vegetation on the southern slopes of Cypress Hills because many lakes immediately south of Cypress Hills are anthropogenic. More recent work based on zonal reconstruction of vegetation from sites across in North America suggests that 13 000 to 14 000 years ago, before the start of the late glacial warming period, Cypress Hills and the Sweet Grass Hills demarcated the meeting point between the western Cordilleran forest and the eastern boreal forest in a thin zonal band adjacent to the Laurentide ice sheet [31]. According to this model, the band of Cordilleran forest extended from west of Cypress Hills to the Rocky Mountains. While it did not extend far into the ice-free corridor of the Rocky Mountains until much later [32], the forest did remain adjacent to the Rocky Mountains in a narrow band following the Rocky Mountains south into Montana. As such, the Cordilleran forest was continuous with site MT1, the only location in the U.S. where Clade B has been found [10]. The band of boreal forest that extended from Cypress Hills is also thought to have been connected to the Black Hills, a site where O. cooperi is common. These bands of forest were transient and replaced by prairie within 2 000 years [33]. While the most recent glacial maximum reached the farthest south, previous glacial maximums could have resulted in forests connecting to Cypress Hills as well, providing earlier corridors [34]. Although sky islands such as Cypress Hills and the Black Hills were once connected, the extent to which Oreohelids used these corridors is uncertain. The vegetation would have been capable of supporting Oreohelids, however, they also require the presence of other abiotic factors, such as calcareous deposits [13]. Limited fossil evidence exists of Oreohelids east of Montana during the most recent forest expansion [35, 36], so they were either present in low densities or absent.
Glacial refugia and colonization
Cypress Hills may have acted as a glacial refugium for at least some Oreohelids during the LGM. It is uncertain if Cypress Hills was habitable during the LGM and there is a lack of consensus in the literature [15, 37]. Some studies suggest it may have been habitable for some species such as lodgepole pine (Pinus contorta) [38, 39], but not other species such as jack pine (Pinus banksiana) [40]. The absence of fossil evidence of Oreohelids in the Great Plains is consistent with Oreohelids already being present in Cypress Hills or the Sweet Grass Hills during the LGM [35, 36]. This does not solve the problem of how the snails colonized Cypress Hills in the first place. Including COI data from [10], the snails of Cypress Hills are paraphyletic and could not have evolved in situ.
Clade X is the only clade in Cypress Hills that is not present on other sky islands to the south or elsewhere in their range and is therefore likely a glacial relict. It is disjunct from its closest relative, Clade E, in northern Wyoming (WY2) over 500 km to the south. Further evidence that Clade X is a glacial relict is the pattern of the haplotype network and distribution of Clades X and B (Fig. 5). The starburst pattern in each is typical of recent population expansions (For more examples, see: [41, 42]). Clade B in Cypress Hills occupies a much smaller proportion of available habitat than Clade X. This could be evidence of Clade B being a later introduction to Cypress Hills. It is likely that Clade B was present in the Rocky Mountains prior to the LGM. Furthermore, the sharing of ITS2 alleles between the mitochondrial groups is indicative of introgression between the two clades. It also shows asymmetry with Cluster 1 containing most of Clade X snails and the presence of two distinct genetic clusters. A second, less likely, possibility is that Clade B was present in Cypress Hills during the LGM and the Rocky Mountains were colonized from the east rather than from the south. The presence of Clade B’ in the Rocky Mountains indicates that Clade B was present in the Rocky Mountains long enough for Clade B’ to diverge. Clade B contains several shared haplotypes in Cypress Hills and the Rocky Mountains, suggesting recent connectivity.
Clades B, C, D, E, and X from our study and [10] are relatively closely related. This species complex likely originated in Montana or Wyoming as most of the diversity occurs in that area, and Oreohelids have been present in Yellowstone Park area for more than 5 million years [29]. Temperature and precipitation fluctuations of the Pleistocene and early Holocene may have allowed colonization of the sky islands via habitat corridors and then isolated each sky island. Ice caps covered many of the sky islands that are currently occupied by Oreohelid land snails, but the distances between them are relatively small and sky islands were connected by suitable habitat for longer periods of time [10, 43]. Similar patterns have been found in glacial relict montane grasshoppers (Melanoplus sp.) in the same area [44, 45].
The restricted distribution of Clade B’ in the Rocky Mountains suggest more recent separation and divergence in situ from Clade B. The divergence between Clades B and B’ is much less than the divergence between any of the other subspecies of O. subrudis [10]. Clade B’ also has reduced ITS2 diversity. The Rocky Mountains foothills contained tundra-like vegetation throughout most of the Wisconsinan and an ice-free corridor throughout the last glacial maximum [32]. Trees did not colonize the majority of the ice-free corridor until approximately 8000 years before present, which is roughly concordant with pollen core data from Harris Lake to the east [22, 32]. Despite extensive surveys, we have not found Oreohelids above 1600 m in the Rocky Mountains. This is the elevation at which snow often remains year-round in southern Alberta and tundra-like, glacial relict vegetation is common.
The ITS2 haplotypes of O. cooperi in Cypress Hills are much more divergent than those found in either Clade B or X, suggesting that O. cooperi may have been in Cypress Hills for a longer period of time. Evidence counter to an early colonization is that the COI haplotypes found in Cypress Hills are shared with snails from both the Judith Mountains and Black Hills. The original source of O. cooperi is likely somewhere within or near Wyoming, which contains O. cooperi, O. pygmaea, and Clade A. These clades form a monophyletic group and likely diverged from the same source population.
Contemporary patterns
The divide between Clades B and X in Cypress Hills is associated with Highway 41. Highways have been demonstrated to be substantial barriers to terrestrial snails [46]. Highway 41 was constructed in the early 1900’s and does not explain the presence of two distinct lineages on either side. The highway may currently reduce gene flow between Clades B and X, but the two clades have been maintained in Cypress Hills since at least the LGM. The niches occupied by both Clades B and X are indistinguishable in Cypress Hills, and these snails have high population densities which may resist invasion from other taxa. Terrestrial snails are known for their ability to rapidly colonize or invade novel ecosystems due to their high reproductive capacity [47]. The spread of the colonizing snails would only stop at the edge of suitable habitat or an occupied niche. Previous work in pipits (Anthus trivialis and A. spinoletta) and buntings (Emberiza citronella and E. hortulana) has shown that long-term species segregation is possible through the combination of factors including habitat differences and interspecific competition [48].
While hybridization between the two mitochondrial clades is evident, it is limited to a narrow area and the introgression of ITS2 alleles is asymmetrical. Many more Clade B snails share ITS2 alleles with Clade X than vice versa. Most hybrids are limited to the contact zone. Oreohelids are simultaneous hermaphrodites, yet the pattern is similar to male-biased dispersal in dioecious organisms [27] suggesting unequal movement between male and female gametes in the contact zone. It could be that these snails seek out mates, but return to their usual resting place to give birth. This would mimic male-biased dispersal, and reduce inbreeding. Another possibility is asymmetrical mating, where snails may behave as males for some mates but females for others. Asymmetrical mating has been demonstrated in land snails including Oreohelids [49, 50] and has been observed in Oreohelids from Cypress Hills (Z. Dempsey, personal observations).
{kind=link}
{kind=link}
{kind=link}