Characterization of enzyme-mediated quinone reaction
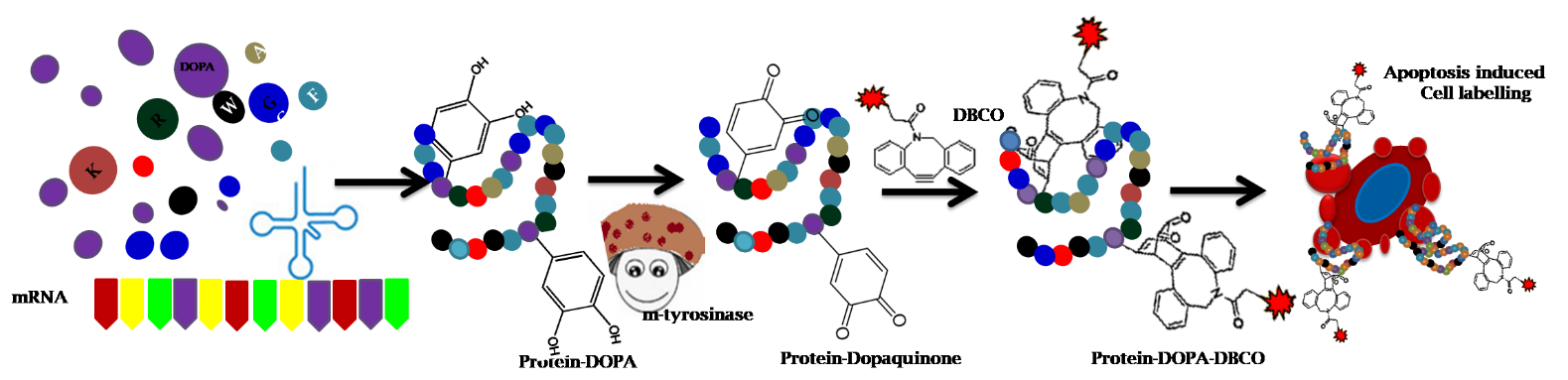
The cycloaddition reaction was carried out using dopaquinone, to confirm the catalytic action of tyrosinase towards the substrate DOPA (Fig. 1C). At first, the pertinence and hurdles of the reaction were examined at a small molecule level (DOPA). Hence, DOPA was treated with 60 U of tyrosinase to produce dopaquinone intermediate in tris buffer pH 6.8, which is further converted into dopachrome within a second (t1/2>1s), by an intramolecular cyclization (Michael- 1, 4 additions) and forms stable melanin pigment that is present in the skin, hair and eyes of all animals [31-32]. As reported by Dukler et al, we also observed a similar absorption spectrum peak corresponding to dopachrome at 475 nm, as a consequence of tyrosinase catalysis (Fig. 1D-1). The conversion of DOPA to dopachrome does not lead to a reduction in the absorbance but leads to a gradual shift in the absorption spectrum [33]. In contrast, a sharp absorbance reduction, a straight line in the UV-Visible spectral region (200-600 nm) was obtained after the tyrosinase treated DOPA conjugated with BCN (Fig. 1D-1), denoting that the reaction is stabilized after the addition of BCN. The absorbance of tyrosinase treated DOPA was compared with BCN conjugated dopaquinone that provided the relative amount of DOPA conjugated with BCN. The conjugation efficiency between dopaquinone and BCN was discovered to be around >90%, which was determined by measuring the difference in the intensity of absorbance maxima at 475 nm before and after BCN conjugation. This data illustrated that the in situ generated quinone through enzymatic catalysis rapidly reacts with the triple bond of the strained alkyne (BCN) via inverse electron demand Diels-Alder cycloaddition, which is sufficient for the generation of DOPA-BCN conjugate. DOPA conjugation was also confirmed by HR-MS analysis with expected molecular mass for DOPA (198.07 m/z) (Fig. 1D-2) and BCN functionalized dopaquinone (355.18 m/z) (Fig. 1D-3). As a positive control experiment, tyrosine was also subjected to tyrosinase treatment and BCN conjugation (Fig. 1A), which also yielded similar results as DOPA (Fig.1B.1-3). These results assured that the formation of dopachrome through intramolecular Michael- 1, 4 additions could be controlled within a short period (10 min) by the addition of BCN.
Biosynthesis of DOPA incorporated protein and purification
To probe the in vivo expression of DOPA incorporated proteins, we selected GFP and anX having different numbers of surface-exposed tyrosine residues. Before proceeding with the protein expression studies, tyrosine residues that are located at the protein’s (anX and GFP) surface was perceived by extricating a 3D model structure from PDB (Fig. 2A-B). Consequently, through the energy minimization process in the protein modeling, it was predicted that anX has six tyrosine residues (Y94, Y133, Y148, Y256, Y257 and Y297) on the surface, out of 11 residues. GFP has eight tyrosine residues including chromophore Y66. Out of these 8 residues, 5 residues (Y92, Y151, Y182, Y200 and Y273) are surface exposed, Y143 is partially surface exposed and Y74 is buried internally (GFP and anX sequence information is provided in the supporting information). Next, to perform the tyrosinase catalysed bio-conjugation reaction on DOPA incorporated proteins, GFP and anX were cloned and the expression was optimized in an E. coli strain JW2581 using 1 mM IPTG [34]. Simultaneously, DOPA was introduced into the proteins (GFP and anX) through the mis-aminoacylation method after depleting the amino acid tyrosine [35]. The expression efficiency of GFP-DOPA and anX-DOPA was compared to native proteins via 12% SDS-PAGE analysis (Fig. 2C). The protein with the substitution of DOPA did not significantly alter the expression of proteins at 37 °C. Further, the protein purification was achieved by the Ni-NTA affinity column and was desalted using AKTA Explorer FPLC (Fig. S2). The incorporation efficiency of DOPA in anX was measured and was compared with wild-type proteins by MALDI-TOF analysis (Fig. 2D). The mass of the GFP increased from 28267 Da to 28396 Da on DOPA incorporation (data not shown), and the mass of anX increased from 35937 Da to 36129 Da, indicating the incorporation efficiency of > 98%.
Structural and Functional Characterization of Modified proteins
To resolve the secondary structure of proteins after DOPA incorporation, the protein samples (GFP and anX) were subjected to CD analysis. Typically, GFP is a beta-sheet protein, showing the positive and negative bands at 195 and 218 nm, respectively whereas anX predominantly has alpha-helix with a negative deflection at around 222 nm and 208 nm (Fig. 2E, and S3). These two proteins have retained secondary structure with a characteristic beta-sheet and alpha-helix pattern after DOPA incorporation. The functional property of these modified proteins viz., GFP-DOPA and anX-DOPA was also evaluated through fluorescent spectroscopic and direct FITC-cell labeling studies, respectively. From these studies, it was demonstrated that GFP-DOPA could maintain its functional property by emitting the orange fluorescence at 525 nm due to the presence of DOPA at the Y66 position of chromophore forming peptide (Fig. S3.B) and anX-DOPA was able to bind to phosphotidyl serine in the lipid membrane of apoptotic cells (Fig. 4C).
Coupling mechanism in protein for cell labeling
To establish a protein-based cell labeling method, the tyrosinase-mediated modification was explored on DOPA incorporated proteins (GFP and anX). In principle, the formation of a quinone from tyrosine by enzymatic and chemical methods is one of the rapid reactions that non-specifically create covalent cross-linking. This is due to the electrophilic quinone that is readily attacked by the proximate nucleophiles especially lysine, histidine and cysteine residues in the protein via Michael type additions [36]. Earlier FREMY'S SALT was tested for the direct conversion of tyrosine to Dopaquinone without the formation of any intermediate. The chemical oxidant-mediated conversion of tyrosine to dopaquinone requires a longer incubation time (20 h) and a high concentration of FREMY’S salt (10M excess) for polymerization, Intra cross-linking of proteins and peptides and conjugation of fluorophore [26-37]. Secondly, FREMY’S SALT is a strong oxidant, hence it could polymerize, aggregate and precipitate the protein. In such a condition, natural enzyme tyrosinase is necessary for reducing the limitations of oxidants like FREMY’S Salt[37]. Usually, chemoselective conjugation for proteins are primed on surface-exposed residues in a suitable reaction condition since it provides high selectivity and accessibility of enzymes towards the least distributed tyrosine residues on the surface. Dopaquinone product formation in proteins was optimized by incubating the DOPA incorporated protein (0.5 mg) with 120 U of tyrosinase in Tris buffer pH 6.8 (Fig. 3A). At this pH, the degree of polymerization and cross-linking that is mediated by a free amino group in the lysine residues is prevented [38]. To substantiate the dopaquinone formation in proteins, a redox staining assay was executed using NBT, for which tyrosinase treated samples were transferred onto PVDF membrane after SDS-PAGE. As a result, purple color owing to the redox cycling of dopaquinone and polymerization (shearing of proteins) was observed on the membrane, which corroborates the conversion of DOPA incorporated protein into quinoprotein by the action of tyrosinase [39] (Fig. 3B.4). Further, residue-specific modification of anX-DOPA was supported by mass spectrometry (MALDI-TOF) with the expected mass shift (36129 Da) (Fig. 2D). Our results reinforced the previous statement on DOPA modification to produce quinone in a single step and it was identified that 1, 2-quinones could quickly react with BCN as described earlier [7]. Therefore, we aimed to functionalize BCN with DOPA incorporated proteins by mild stirring for 2 h at RT. After dialysis, BCN functionalized anX-DOPA was confirmed by SDS-PAGE (Fig. 3B.3) and MALDI-TOF analysis with anticipated mass shift (37071 Da) (Fig. 3B.1), representing that five molecules of BCN would have reacted with enzyme catalysed-anX-DOPA. The suitability of the reaction for residue-specific labeling was inspected and were allowed to conduct a SPOCQ on quino-proteins. The anX-DOPA was coupled with a strained alkyne namely fluorophore DBCO by the action of the enzyme tyrosinase and analyzed by SDS-PAGE (Fig. S4) and MALDI-TOF analysis with a mass shift (40808 Da) indicating that five molecules of DBCO is conjugated with surface-exposed DOPA residues of anX-DOPA (Fig. 3B.2). Moreover, tyrosinase catalyzed anX-DOPA-DBCO conjugate exhibited emission spectrum at 595 nm after dialysis (10 kDa), which shows that the anX-DOPA was tagged with the fluorophore (DBCO) (Fig. 3C). In contrast, anX-DOPA-DBCO was also prepared as a control experiment without tyrosinase, which did not exhibit fluorescence when it was exposed to UV light after the dialysis (Fig. 3C.1-3). It denotes the importance of tyrosinase for residue-specific protein modification and its key role in labeling experiments. The generality of this method can be applied to any proteins containing fewer numbers of surface-exposed tyrosine residues. To test the application of fluorogenically labeled quino-protein, we employed the single-step strain-promoted oxidation-controlled click reactions for cell labeling and imaging experiments. Hence, the fluorogenically labeled anX-DOPA was used for apoptosis recognition.
Fluorescent microscopy and flow cytometry analysis of apoptotic EA.hy926 cells
During programmed cell death or apoptosis, the cell loses the phospholipid asymmetry of the plasma membrane, which reshuffles phosphatidylserine (PS) from its usual position [40]. In the presence of Ca2+, the protein anX exclusively and sturdily binds with the PS molecule. Therefore, commercially available fluorescent tags such as FITC, Oregon-Green, or Alexa Fluor 488, succinimidyl esters were linked with anX by chemical coupling methods and they were extensively applied to probe apoptosis incidence. However, the classical coupling reactions cannot be controlled due to the presence of the free amino group in the lysine residues, which are mostly surface exposed and exist predominantly in or near the Phosphatidylserine binding pocket.
The consequent conjugation of Annexin with FITC would probably result in fluorescence quenching and yield a heterogeneous mixture [41-42]. The added advantage of genetic code engineering is to develop a highly controlled and homogenously labeled fluorescent anX conjugate without altering its functional property. To evaluate the efficacy of anX-DOPA-DBCO for cell labeling, apoptotic studies were done in EA.hy926 cells. Apoptosis was induced in EA.hy926 cells by treating with Doxo, and the apoptotic changes were observed using phase-contrast microscopy. Cells with flattened and disruptive membrane morphology were observed at the concentration of 50 µg/mL Doxo (Fig. 3D). The PS-binding activity of anX-DOPA-DBCO was analyzed on apoptotic EA.hy926 cells through flow cytometry. It was compared with the commonly accessible anX-FITC. 0.5 µg/mL concentration of anX-FITC and anX-DOPA-DBCO were used to stain the apoptotic cells (Fig. 4B-C) and it was compared to the cells without apoptosis induction (Fig. 4A). The cells treated with anX-DOPA-DBCO responded well for apoptosis and it was more or less similar to anX-FITC, which indicated that apoptotic labeling using anX-DOPA-DBCO resulted in PS binding even after protein modification (Fig. 4B-D).
Moreover, the advantage of using DBCO (orange fluorophore) over FITC is that it has low quantum yield but high emission maxima, lower light scattering, and reduced autofluorescence [43]. FITC (shorter wavelength) with green spectral window is much more energetic than DBCO (longer wavelength) with red spectral window, as a result, shorter wavelength fluorophores including FITC, DAPI, Alexa-488 has potential photo-bleaching, lesser photo-stability and pH sensitivity in comparison with the light of longer wavelength orange and red fluorophores (DBCO) [44]. From our results, we postulated that tyrosinase-mediated modification in anX-DOPA did not alter the function of proteins, especially for apoptosis detection. Though it is too preliminary, however, it could pave a way to extend the single-step strain promoted oxidation-controlled click reactions via expanding the genetic code to the other caspase pathway proteins such as Bax and Bcl-XL, which are involved in the early and late events of apoptosis.
{kind=link}