The BLTS-humanized mouse model supports functional human innate and adaptive immune cells in the blood and the human lymphoid tissues.
We previously reported human immune cell development in the peripheral blood and the human primary and secondary lymphoid tissue grafts in the BLTS-humanized mouse model [22]. Here, we demonstrate the reconstitution of additional human immune cell types in peripheral blood and human primary (thymus) and secondary (spleen) lymphoid tissue grafts. Specifically, we demonstrate reconstitution with NK cells, CD4+ and CD8+ T cells, monocytes, memory B cells, plasmablasts, and pan-T cells, pan-B cells, and macrophages as previously demonstrated (Supplementary Figure 1-3) [19]. Here, we also demonstrate the functionality of human-antigen presenting cells in the BLTS-humanized mouse model (Supplementary Figure 4A, 4B). Bone marrow-derived hematopoietic stem cells in the BLTS-humanized mice can differentiate into type-1 and type-2-polarized dendritic cells (DCs) (Supplementary Figure 4A). Type-1 DCs secrete the T-helper 1 (Th1)-driving cytokine (IL-12p70) upon stimulation with physiological ligand (CD40L) (Supplementary Figure 4A). Human CD163+ splenic macrophages from BLTS-humanized mice phagocytized bacteria (pHrodo™ Red E. coli BioParticles) upon co-culture (Supplementary Figure 4B). Furthermore, we demonstrate the functionality of T cells in the BLTS-humanized mouse model. We show robust splenic T cell response (type-1 cytokine, IFNg) to physiological stimulation (CD3 and CD28) that was comparable to the cytokine (IFNg) response by T cells from human PBMCs (Supplementary Figure 4C, 4D).
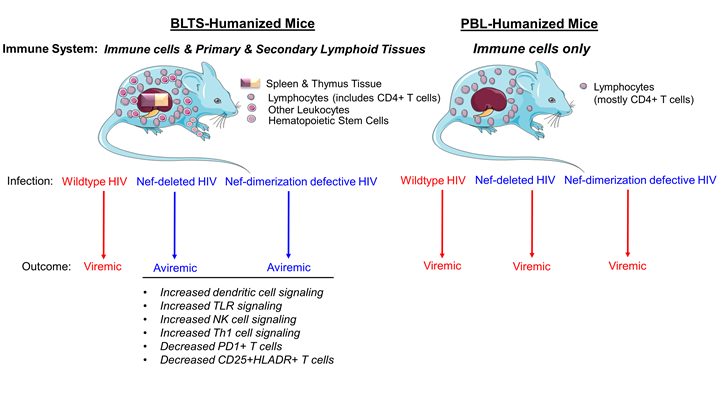
The human immune system in the BLTS mouse model abrogates nef-deleted HIV viremia.
We previously reported that BLTS-humanized mice support HIV replication and associated immunopathogenesis [19]. We demonstrate that human macrophages (CD163+ splenic cells) and CD4+ T cells facilitate HIV replication (Supplementary Figure 5) [19]. Therefore, BLTS-humanized mice provide a robust in vivo model for investigating Nef-immune system interactions in HIV infection. Nef enhances HIV infectivity and replication in some cell lines [39-41]. Nef also enhances HIV replication in human PBL-derived activated-primary CD4+ T cells in a viral input-dependent manner, with the Nef effect lost at a high viral-input dose [42]. Here we show that nef-deleted HIV (NL4-3 strain) exhibits reduced replication compared to wild-type HIV in the TZM-bl cell line (Figure 1A). However, nef-deleted HIV and wild-type HIV exhibit negligible differences in viral replication at high viral input in the activated-primary CD4+ T cell culture model. (Figure 1B). The in vivo equivalent of the activated-primary CD4+ T cell culture model is the PBL-humanized mouse model [17, 43]. Using the PBL-humanized mouse model, we demonstrate that nef-deleted HIV and wild-type HIV exhibit similar infectivity and replication in the blood and lymphoid tissues (Figure 1C, 1D, 1E), which is consistent with previous studies [44]. On the contrary, inoculation of BLTS-humanized mice with nef-deleted HIV and wild-type HIV resulted in divergent outcomes, with viremia in wild-type HIV inoculated mice and no viremia in nef-deleted HIV-inoculated mice (Figure 1C, 1F). We inoculated BLTS-humanized mice with the same high dose of nef-deleted HIV and wild-type HIV stocks used to infect PBL-humanized mice; therefore, the nef deleted HIV inoculum is infectious and replication-competent (Figure 1C). These observations demonstrate that Nef overrides host immune control of HIV-1 replication in the BLTS-humanized mouse model.
HIV transmission in humans results in a cytokine burst (including IFNg, IL-10, and TNFa) within two weeks of exposure [45], which stimulates immune cells to initiate an anti-viral immune response [46]. Inoculation of BLTS-humanized mice with nef-deleted HIV induces a modest increase in the secretion of type 1 cytokines (IFNg, IL-2, TNFa, IL-12p70) and type 2 cytokines (IL-10, IL-6, IL-8, IL-13) as compared to the highly viremic wild-type HIV (Supplementary Figure 6). The cytokine burst in wild-type HIV infection does not eliminate the virus or reduce viremia to undetectable levels; instead, it may fuel immune dysregulation [45]. The resulting chronic HIV infection induces progressive T cell-immune activation and checkpoint inhibitor expression [45, 47, 48], inadequate anti-viral T cell responses [45, 47, 48], and immune dysregulation [49, 50].
Nef dimerization defect abrogates HIV viremia and associated immunodeficiency in the BLTS-humanized mouse model.
Previous X-ray crystallography studies of HIV-1 Nef proteins either alone or in complexes with host cell kinase regulatory domains revealed that Nef forms homodimers (reviewed in Staudt et al., 2020). Comparison of these structures identified several residues common to these Nef dimer interfaces, including L112, Y115, and F121. Mutagenesis of these residues prevents homodimerization of recombinant Nef in vitro and reduces Nef dimer formation in a cell-based bimolecular fluorescence complementation (BiFC) assay. [29, 51]. Based on the BiFC assay, the relative disruption of dimerization in the Nef mutants is: L112D+Y115D double mutant > Y115D > F121A [29, 32]. Previous studies showed that these dimerization-defective Nef mutants also reduce HIV replication [29, 32] and abrogate HIV-induced impairment of host defense [33] in tissue culture models, implicating Nef dimers in signaling mechanisms by which HIV-Nef promotes viremia and immune dysregulation [29]. Interestingly, we show that the nef-dimerization defective HIV strains, Y115D, and L112D+Y115D are replication-competent in PBL-humanized mice and the HSC-humanized mice (Supplementary Figure 7), respectively. However, BLTS-humanized mice inoculated with the nef dimerization-defective HIV strains Y115D and Y115D + L112D were aviremic (Figure 2, Supplementary Figure 8) and demonstrated reduced HIV-infected T cells in human spleen xenografts when compared to wild-type HIV infection (Figure 3A, 3B, Supplementary Figure 9). Additionally, mice inoculated with HIV carrying the Nef-F121A mutant were partly viremic (67% of mice) (Supplementary Figure 8). BLTS-humanized mice inoculated with nef-deleted HIV (NL4-3 strain) were predominately aviremic (75% of the mice) and had low levels of HIV-infected T cells in the human spleen xenografts (Figure 2, 3A, 3B). Additionally, nef-defective HIV (DNef and the dimerization-defective mutant, Y115D) and mock-inoculated BLTS-humanized mice exhibited similar CD4+/CD8+ T cell ratios, whereas wild-type HIV-infected BLTS-humanized mice exhibited immunodeficiency (reduced CD4+/CD8+ T cell ratio; CD4+ T cell loss) (Figure 4A, 4B).
Nef dimerization defect attenuates HIV-induced immune dysregulation in the BLTS-humanized mouse model.
Many studies have shown that Nef impairs the anti-viral immune response to HIV infection [52]. Furthermore, several lines of evidence demonstrate that Nef-dimerization defective mutants and Nef inhibitors abrogate HIV-induced immune dysregulation [8, 37]. We observed that Nef dimerization defect (Y115D mutant) attenuates HIV-induced T cell-immune activation (elevated CD25+ and HLA-DR+ levels) (Figure 5, Supplementary Figure 10A) and checkpoint inhibitor expression (elevated PD1+ levels) (Figure 6, Supplementary Figure 10B) in the blood and human spleen. Consistent with the role of Nef in promoting HIV-induced immune dysregulation, complete deletion of the nef gene in the HIV genome attenuates HIV-induced T cell-immune activation markers (elevated CD25+ and HLA-DR+ levels) and checkpoint inhibitor expression (high PD1+ levels) in the blood and the human spleen in BLTS-humanized mice (Figure 5-6, Supplementary Figure 10).
The current consensus posits that chronic HIV infection in humans results from an inadequate antiviral-Th1 immune response, which is associated with T cell-checkpoint inhibitor expression [47], systemic immune inflammation (i.e., elevated levels of CXCL13) [53], B cell hyper-activation and dysregulation [27, 49], and reduced levels of anti-viral factors (i.e., CXCL12) [54]. Previous studies also show that HIV Nef dimers are required to activate non-receptor tyrosine kinases of the Src and Tec families to enhance viral replication (Staudt et al. JBC 2020) and are also linked to downregulation of MHC-I and CD4 receptor on T cells [29, 55]. Additionally, Nef promotes B cell hyper-activation and dysregulation; this pathogenic effect has been associated with T cell interaction [27, 49]. Dysregulated immune activation and signaling by Nef are presumed to drive chronic HIV infection and progression to AIDS [56]. Gene expression analysis demonstrates that wild-type HIV infection in BLTS-humanized mice induces systemic inflammation marker (elevated CXCL13), B cell activation markers (elevated CD19, CD79, PAX5, and BLNK), and reduced levels of anti-viral factors (CXCL12, PML/TRIM19) in the human spleen xenograft (Figure 7A). Wild-type HIV infection also induced the expression of the non-receptor tyrosine kinase, ABL1, in the human spleen xenografts of BLTS-humanized mice (Figure 7A). ABL1 plays a role in T cell receptor signaling and cytoskeletal rearrangement, facilitating HIV entry into the host cell [57]. ABL1 also enhances HIV-1 replication by activating RNA pol-II [58]. Comparative analysis of gene expression in BLTS- humanized mice inoculated with wild-type HIV and nef-deleted HIV showed reduced expression of the non-receptor tyrosine kinase, SYK, reduced systemic inflammation (CXCL13, HLA-DR), decreased B cell activation markers (CD19, AICDA), and elevated levels of anti-viral factors (CXCL12, LIF) in the human spleen xenografts in nef-deleted HIV infection compared to wild-type HIV infection (Figure 7B). A similar analysis of the human spleen xenograft revealed reduced systemic inflammation (CXCL13); elevated levels of anti-viral factors (CXCL12, IFIT2); upregulation of genes involved in complement activation (C1Q); increased expression of RAG1/2 for the development of a diverse repertoire of immunoglobulins; and elevated levels of the MHC-related molecule CD1A in BLTS-humanized mice challenged with the nef-dimerization mutant (Y115D) as compared to mice challenged with wild-type HIV (Figure 7C).
Consistent with the gene expression profile in the human spleen in BLTS-humanized mice, Ingenuity pathway analysis (IPA) demonstrates elevated systemic immune activation signaling (Th1 pathway, Neuroinflammatory pathway, Induction of apoptosis by HIV) and B cell signaling (PI3K signaling, FcgRIIB signaling, B cell receptor signaling) in HIV infection (Figure 8A). It also revealed reduced anti-viral immune signaling, specifically Toll-like receptor (TLR) signaling, HMGB1 signaling, NK cell signaling, and RIG1-like receptors, all of which are downregulated in wild-type HIV infection compared to mock (Figure 8A). Additionally, IPA demonstrates elevated anti-viral immune signaling (Toll-like receptor signaling, HMGB1 signaling, complement system signaling, NK cell signaling, DC maturation, and NK-DC crosstalk, RIG1-like receptors and pattern recognition receptors (PRR) signaling) and enhanced Th1 signaling and decreased B cell signaling (PI3K signaling, FcgRIIB signaling, B cell receptor signaling) in nef-defective HIV infection cohorts (DNef and dimerization-defective Nef-Y115D) compared to wild-type HIV cohorts (Figure 8B, 8C). This suggests that components of the innate immune system in the human spleen xenografts such as complement system, PRRs, HMGB1, NK cells, dendritic cells, and macrophages are playing an essential role in controlling viremia in nef-defective HIV infected humanized mice. Furthermore, deletion or mutation of nef abrogates the ability of HIV to interact with host proteins to downregulate MHC class-I, which allows CTLs to recognize infected cells in the context of MHC class-I and induce cytotoxicity to control viremia in BLTS-humanized mice.
Several studies have demonstrated that HIV Nef promote NFAT-dependent signaling, which in turn promote HIV replication [59-61]. IPA of human spleen xenograft-gene expression demonstrates elevated NFAT-dependent signaling in the wild-type HIV group compared to the mock group (Figure 8A). Importantly, IPA of human spleen xenograft-gene expression demonstrates reduced NFAT-dependent signaling in nef-deleted HIV group compared to wild-type HIV group (Figure 8B), which suggests that Nef is modulating immune signaling in the human spleen xenograft.
{kind=link}