iPSCs
Morphological examination of cell cultures revealed remarkable differences between control (Ctrl) and RTD iPSCs. Phase contrast microscopic analysis demonstrates reduced ability to form colonies shown by RTD cells, compared to healthy iPSC cultures, which instead showed regular margins of the colonies and stable cell-cell contacts as expected [9]. Notwithstanding general common features, RTD cells displayed patient-specific morphological abnormalities, in that SLC52A2mut (p.S52F; p.L312P) iPSCs were totally disaggregated, while Pt.1 (SLC52A2mut p.S52F; p.G149S), Pt.2 (SLC52A2mut p.Arg169Cys; p.Leu344Alafs100) and Pt.3 (SLC52A2mut p.Arg169Cys; p.Trp198Ter) form small, irregular colonies. Such alterations related to altered cell morphology, suggesting cell suffering and/or ongoing death process.
Based on these observations and previous findings [9, 10, 11] suggesting impaired survival of RTD cells, we addressed the putative involvement of apoptotic process in RTD patients’ iPSCs. Morphological details of RTD iPSCs were investigated by scanning electron microscopy (SEM), revealing abundant membrane blebbing in RTD iPSCs; on the contrary, Ctrl iPSCs showed a smooth surface (Fig. 1). Additionally, spherical vesicles, ranging from 1 to 5 µm, which are reminiscent of apoptotic bodies, were consistently found to accumulate on apical cell surface in all RTD patients’ cultures.
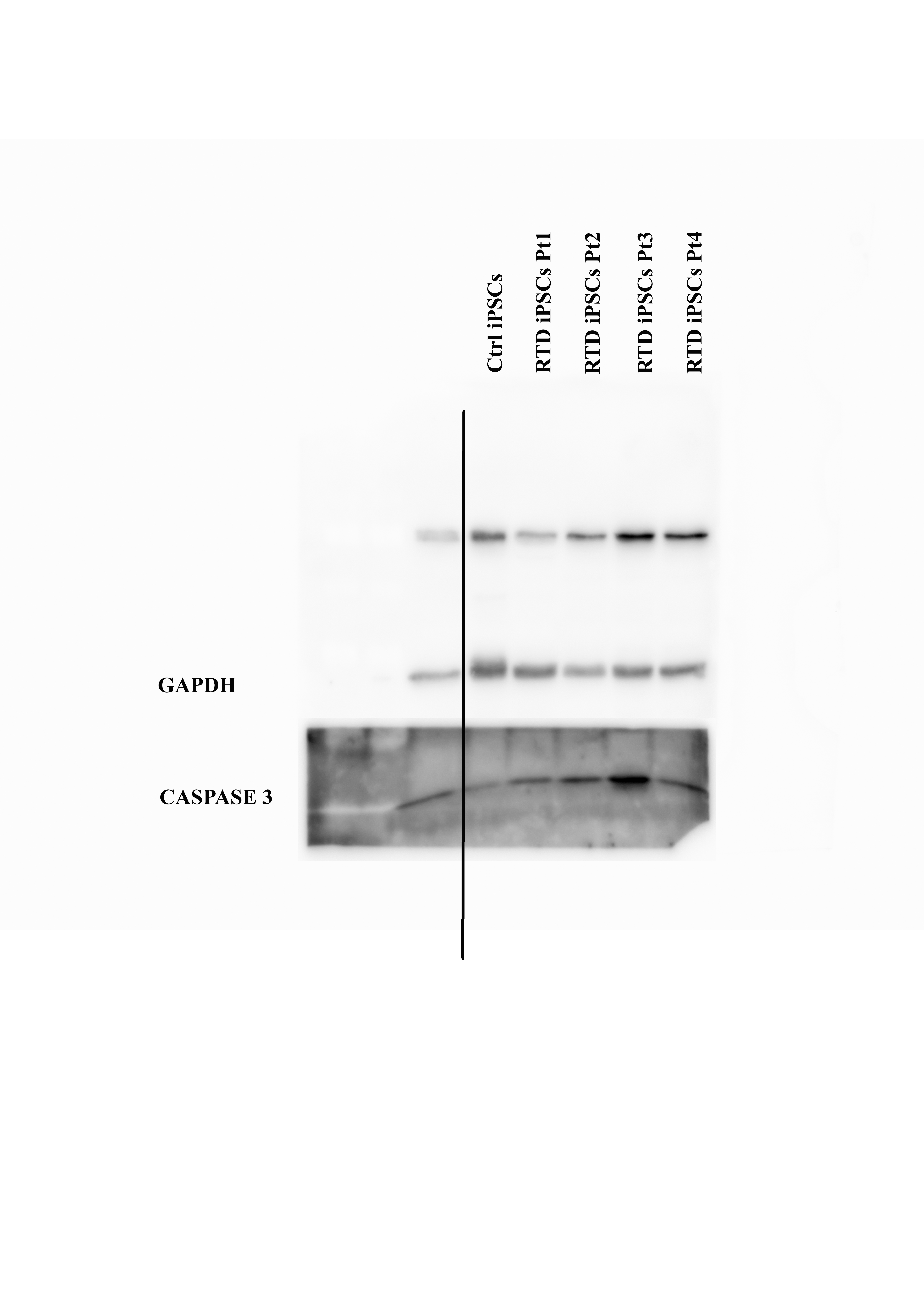
We were then prompted to investigate the apoptotic pathway, particularly caspase activation, studying Caspase 3 immunoreactivity in RTD iPSCs, as compared to controls. Figure 2A displays representative confocal images of Caspase 3 immunofluorescent localization, providing evidence of an abnormally high number of positive cells in RTD cultures. We also observed a heterogeneous intracellular distribution of the apoptotic marker, which was found in the peripheral cytoplasm, next to the plasma membrane. Notably, Caspase 3 positive cells often displayed morphological alterations typical of the apoptotic process, including blebs, apoptotic bodies, and nuclear polymorphism/fragmentation (Fig. 2B). When quantitatively evaluated, Caspase 3 signal intensity proved significantly higher in iPSCs of all RTD patients, as compared with Ctrl cells. However, individual-based differences in the abundance of Caspase 3 were also detected. Indeed, Pt.1 cells display the highest fluorescence intensity levels, while Pt.3 cells were the least immunoreactive (Fig. 2D). These data were consistent with WB findings (Fig. 2E).
We performed TUNEL assay to evaluate apoptotic cell death. Confocal microscopic images showed the presence of a greater level of the nuclear fluorescent signal in RTD iPSCs, compared to Ctrl cells (Fig. 2C).
iPSC-derived MNs
To study cell death process in the cell type mostly affected by RTD, we differentiated iPSCs into MNs and examined them by SEM. Patient-derived MNs showed dramatically altered morphological features, including evaginations or membrane blebbing, and cell shrinkage, hallmarks of apoptotic process (Fig. 3). Moreover, while Ctrl cells appeared firmly adherent to the substrate and formed an intricate network of cell bodies and neurites, RTD cells were often loosely juxtaposed to each other and to the underlying substrate (Matrigel), and extended shorter neurites, consistent with previous studies [8, 11]. Filopodia-like fine structures also emerged from RTD cell bodies.
To characterize the cell death process in RTD MNs, we performed Caspase 3 IF (Fig. 4A) and TUNEL assays (Fig. 4B) on healthy and diseased MNs. Confocal images showed significantly increased percentage of Caspase 3 positive cells in RTD MNs, as compared to controls (Fig. 4A, C).
Double IF, using anti-β III Tub also showed a decreased signal in MNs derived from Pt.1 iPSCs. Apoptotic bodies and nuclei fragmentation were readily detected in MNs from all RTD patients (Fig. 4, A’ column). Consistently, TUNEL assays revealed the presence of a remarkably increased number of apoptotic nuclei in RTD MN, as compared to Ctrl cells (Fig. 4B-D).
All the described alterations related to the activation of the apoptotic pathway, were consistently more severe in Pt.1 MNs, though statistically significant in all RTD differentiated cells.
To ascertain caspase involvement in apoptotic cell death in RTD, thereby excluding caspase-independent pathways, we addressed possible implication of AIF, based on the notion that this molecule is also supposed to be released by dysfunctional mitochondria. Specifically, IF labeling was used to assess changes in AIF expression and localization in RTD MNs. Triple IF, using anti-AIF, anti-mitochondria antibody (MTC02) and anti-βIII Tubulin, showed overall co-localization of AIF/MTCO2 in RTD, as well as in Ctrl MNs (Fig. 5). Interestingly, little or no nuclear AIF localization was detected in any cell.
The series of irreversible events triggered by activation of the apoptotic pathway include chromatin condensation accompanied by DNA degradation. FIB/SEM analyses allowed us to study the nuclear and cytoplasmic fine morphology of Ctrl and RTD MNs (Fig. 6). The latter were characterized by often lobed and even polymorphic nuclei, surrounded by an indented nuclear envelope. Such ultrastructural features remarkably differed from that observed in Ctrl cells, which instead showed rounded-shape nuclei and a regularly arranged nuclear envelope. Furthermore, intranuclear inhomogeneities were observed, possibly corresponding to chromatin fragmentation (Fig. 6A, arrow). Ultrastructural analysis of RTD MNs also confirmed mitochondrial abnormalities associated with RTD phenotype. Patients’ cells contained few mitochondria, often immature or degenerated, with the presence of few, fragmented or swollen cristae, and with an altered organization of the inner mitochondrial membrane (Fig. 6B-C). To make a semi-quantitative evaluation of RTD-associated phenotype severity, we classified mitochondrial morphology as healthy, mildly affected and disrupted, based on as the following features: cristae fragmentation and outer membrane ruptures (Fig. 6F). Statistical analyses confirmed the presence of a significantly greater number of either severely or mildly damaged organelles in RTD than in Ctrl cells.
Considering the mitochondrial damage observed in RTD MNs carrying SLC52A2 mutations, resulting in defective RFVT3 transporter, we hypothesized that the morphological alterations could be related to the presence and function of RFVT3 protein in these organelles. To test this hypothesis, we performed double IF using anti-RFVT3 antibody and anti-mitochondria antibody (MTC02). Interestingly, RFVT3 displayed a granular immune distribution, which in patients tends to form clusters with abnormal distribution. Moreover, the signal is partially co-localizing with the mitochondrial marker in control and patient-derived neurons. Representative images obtained at the confocal microscope are shown in Fig. 7.
{kind=link}