JH enhances the weight and thickness of silkworm cocoon
To identify the function of JHA in improving silk protein production in silkworms, we performed individual experiments on silkworm larvae. After JHA treatment for 24 h, we found that the application of JHA significantly extended the fifth larval instar of silkworm. JHA treatment extended the B. mori fifth instar from 8 to 11 days (Fig.S1). We further measured the effect of JHA on the weight and thickness of the cocoon. The results showed that JHA enhanced the weight of silkworm cocoons, especially those of male silkworm (Fig. 1A). In addition, we found that JHA treatment significantly increased the thickness of silkworm cocoons, both in males and females (Fig. 1B-C). Taken together, these results showed that the application of JH extended the larval period, increased the weight and thickness of the cocoon, and further enhanced silk production in silkworms.
Characteristics of m 6 A modification in the transcriptome of the B. mori posterior silk gland
To investigate the characteristics of m6A modification in the PSG of silkworm, we performed m6A methylation sequencing to identify specific locations of m6A sites in the mRNA of PSG (Fig. 2A). Our results showed that m6A modification was mainly enriched in exons and 3’ UTRs, with particularly high levels of m6A peaks observed around stop codons (Fig. 2B-C). Through the analysis of consensus motifs, we observed that the GGAC motif was enriched within m6A sites (Fig. 2D). The motif has been frequently found in other studies on humans, mice, and fruit flies. To confirm the role of m6A in the expression of genes, we further analyzed the relationship between the expression level, represented by FPKM, and the m6A ratio of m6A-containing genes. The results showed that the expression levels of m6A-containing genes had a negative relationship with the m6A modification levels of m6A-containing genes (Fig. 2E). Collectively, the m6A sequencing data provided clear evidence of the distribution features and consensus motifs of m6A in the mRNA of silkworm PSG. These results also suggested that m6A modification in the mRNA of silkworm PSG tends to regulate gene expression in a negative manner.
m6A-containing genes involved in several important biological pathways
m6A modification is involved in various gene expression regulation and biological processes [30]. To investigate the role of m6A in silkworm PSG, we performed Kyoto Encyclopedia of Genes and Genomes and GO enrichment analysis on high-confidence m6A-containing genes. From the m6A-seq data, we identified 4962 high-confidence m6A-containing genes. Seventy-four m6A-containing genes are involved in the spliceosome and are critical for mRNA alternative splicing. Forty-four and 37 m6A-containing genes were enriched in the mRNA surveillance pathway and RNA degradation, respectively (Fig. 3A). These pathways are important for the stability of mRNA. The results showed that m6A-containing genes are involved in regulating the stability, splicing and translation of mRNA.
Furthermore, GO enrichment analysis revealed that 244 and 176 m6A-containing genes were enriched in nucleic acid binding and nucleotide binding, respectively. Compared to molecular function terms, biological process terms, including gene expression, nucleic acid metabolic process, and RNA metabolic process, were enriched. Additionally, 277 m6A-containing genes were enriched in the nucleus in the cellular component category (Fig. 3B). These results indicated that m6A is closely related to nucleic acid binding and metabolic processes. Taken together, our findings suggest that m6A plays a critical role in influencing RNA fate and regulates gene expression in the PSG of B. mori.
JH regulates the m6A modification and expression level of genes in silkworm PSGs
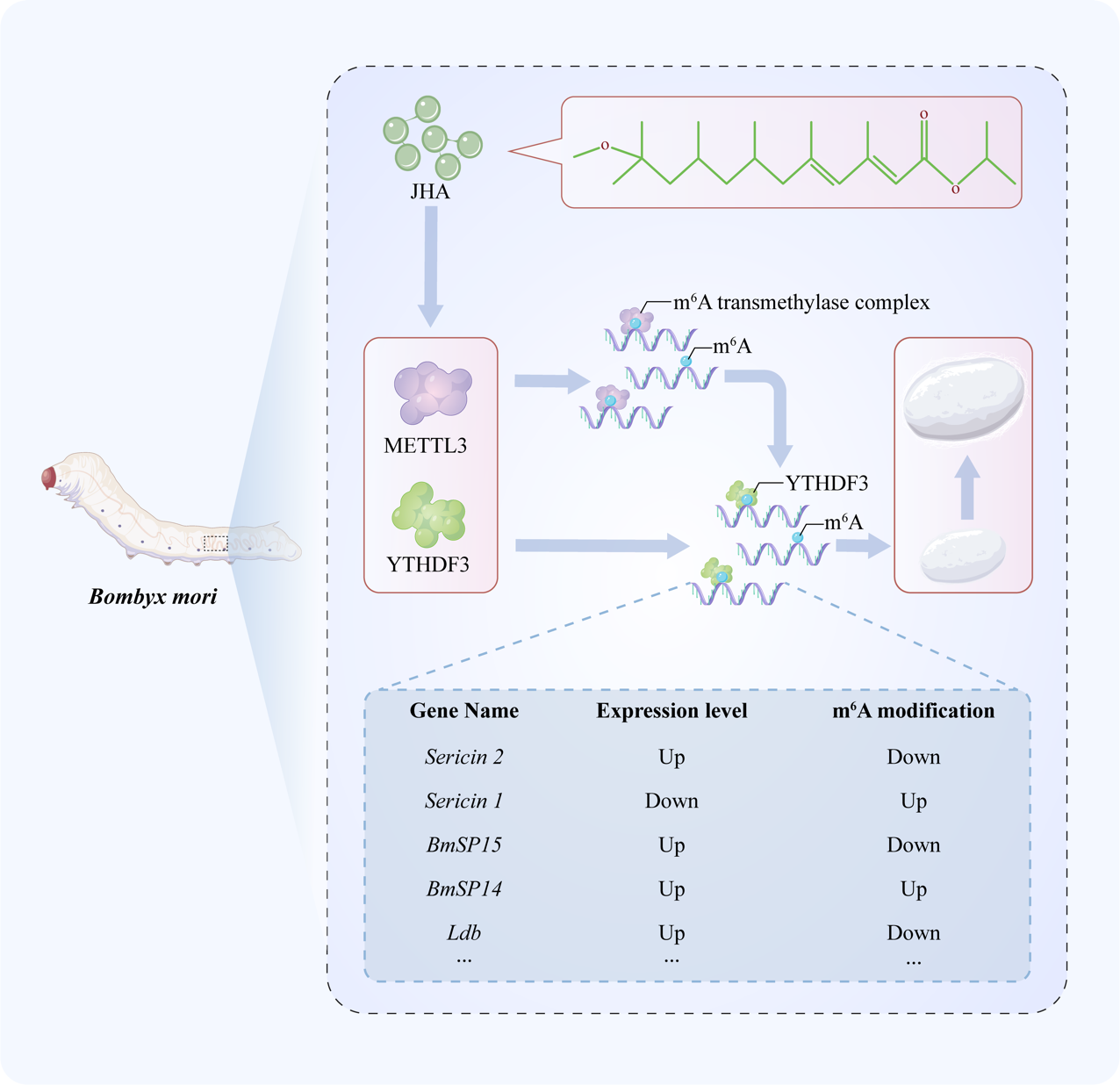
To investigate the relationship between JHA and m6A modification, we performed transcriptome sequencing of multiple silkworm tissues after JHA treatment. Our findings indicated that the expression of m6A-related genes was significantly changed by JHA treatment in PSG compared to other tissues (Fig.S2A). Western blotting also showed that JHA treatment significantly induced the protein expression of METTL3 and YTHDF3 in silkworm PSGs (Fig.S2B). Additionally, we further found that JHA treatment significantly increased the m6A abundance of RNA in the PSG (Fig.S2C). The results indicated that JHA treatment affects the expression of METTL3 and YTHDF3, thereby regulating the m6A abundance of RNA in the PSG of silkworms.
We further performed m6A sequencing of PSGs after JHA treatment. By transcriptome and m6A sequencing data conjointly, we found 3795 differentially m6A-modified genes (DMGs) and 3961 differentially expressed genes (DEGs) in m6A-seq and RNA-seq of silkworm PSG, respectively. DMGs and DEGs were mapped on B. mori chromosomes, and we found that the density of m6A modifications, different m6A modifications, and gene density on chromosomes were highly consistent with each other. In most B. mori chromosomes, downregulated and upregulated DEGs were distributed uniformly. However, the downregulated DEGs were enriched significantly at the telomeres of chromosomes 27 (Fig. 4A). Moreover, 261 downregulated and 222 upregulated genes had upregulated m6A modification levels. A total of 438 and 614 genes with downregulated m6A modification levels were downregulated and upregulated, respectively. These results revealed that JHA widely regulates gene expression and mainly downregulates gene expression by regulating m6A modification of mRNA. Notably, 2304 DEGs were not DMGs, and 1455 DMGs were not DEGs (Fig. 4B-C). Comparing m6A peak lengths between the control and JHA treatment groups revealed that JHA treatment significantly decreased the length of mRNA, which was modified by m6A (Fig. 4D). Although statistical analysis of the number of m6A peaks in each gene, we found that most genes had one or two m6A peaks. JHA treatment decreased the proportion of genes with one or two m6A peaks and increased the number of genes with three m6A peaks (Fig. 4E). These results showed that JH affects the density and distribution of m6A modifications, thereby regulating m6A-containing gene expression.
Enrichment analysis of differentially m6A-modified DEGs after JHA treatment
To investigate the effects of JHA treatment on silkworm PSGs, we performed GO and KEGG enrichment analyses of genes with different expression levels and m6A peaks in RNA-seq and m6A-seq data. We found DEGs with significantly different m6A modifications after JHA treatment. To uncover the function of these genes, we performed GO enrichment analysis. Most of the genes were enriched in molecular function terms related to binding activity, especially nucleic acid binding activity. Two cellular component terms, nucleus and protein-containing complex, as well as five biological process terms, gene expression and biological regulation, were enriched in the GO analysis (Fig.S3). The results further confirmed that m6A modification affected the fate of nucleic acids by regulating nucleic acid binding and nuclear components, thereby regulating gene expression and biological processes. KEGG enrichment analysis of these 547 genes found that 8 pathways, including autophagy, Hippo signaling pathway, MAPK signaling pathway, mTOR signaling pathway, nucleocytoplasmic transport, protein processing in endoplasmic reticulum, spliceosome, and Wnt signaling pathway, were highly enriched (Fig. 5). In total, 61 of 72 DEGs in these 8 pathways were downregulated after JHA treatment. All DEGs were downregulated in 4 pathways, including the Hippo signaling pathway, nucleocytoplasmic transport, spliceosome, and protein processing in the endoplasmic reticulum. The results suggested JHA affects multiple biological processes in PSGs through m6A modification.
JHA treatment regulated the m6A modification and expression level of silk-associated genes
To better understand the effect of JHA on silk protein synthesis in PSGs, we conducted an analysis of the expression changes in silk-associated genes, which are critical for silk protein synthesis, transport, and silk fiber formation (Table S1)[31]. These silk-associated genes were grouped into three functional categories: protease inhibitors, silk proteins, and transcription factors. Among these silk-associated genes, 32 genes were identified in transcriptome sequencing data (Fig. 6A), and 16 genes were significantly differentially expressed (Fig.S4). In addition, 6 of the 35 silk-associated genes showed significant changes in m6A modification (Fig. 6B-F), suggesting that these genes are targets of m6A modification. Notably, m6A modification was mainly concentrated in the 3’ UTRs and CDS regions of these 6 silk-associated genes. Furthermore, we observed that JHA resulted in different m6A modification changes for different gene transcripts. JHA significantly increased the m6A modification levels of seroin 1, proteasome inhibitor PI31, and BmSPI4. However, the m6A modification levels of sericin 2, BmSPI5, and Ldb were downregulated by JHA. Taken together, these results showed that JHA has a profound impact on regulating the expression of silk-associated genes by altering m6A modification.
Knocking down METTL3 regulated the expression level of silk-associated genes
To investigate the role of m6A in the expression of silk-associated genes, we further knocked down the expression of METTL3 in embryonic silkworms. Through transcriptome sequencing data analysis, we identified 3561 DEGs after METTL3 knockdown, and 1359 DEGs were m6A-containing genes. The m6A sequencing data in this study revealed that multiple genes in the JH signaling pathway are also m6A-methylated (Table S2). Knocking down METTL3 widely affected the expression of many genes in the JH signaling pathway, including Juvenile hormone esterase 1 (JHE1), Juvenile hormone epoxide hydrolase 2 (JHEH2), Juvenile hormone epoxide hydrolase 3 (JHEH3), Farnesyl pyrophosphate syntase (FPS), etc (Table S3). We further identified 32 silk-associated genes in the RNA-seq of silkworm embryos (Fig. 7A). In total, 11 silk-associated genes were DEGs (Fig. 7B). Interestingly, 5 DEGs, including BmSPI4, BmSPI5, sericin2, Ldb, and seroin1, had significantly different m6A modification levels after JHA treatment in the PSG (Table S4). Taken together, the results suggested that the expression of BmSPI4, BmSPI5, sericin2, and seroin1 is regulated by m6A modification.
JHA regulated the expression of BmSPI4 and BmSPI5 via m 6 A modification
To investigate whether JH directly regulates silk-associated genes, we treated BmN cells with JHA for 2 h, 4 h, 6 h, 8 h, 12 h, and 24 h (Fig. 8A, Fig S5). The qPCR results suggested that JHA significantly increased the expression of METTL3 and YTHDF3 (Fig. 8A), and the results were confirmed by western blotting analysis (Fig. 8B). We also found that BmSPI4 and BmSPI5 were induced by JHA in BmN cells at 12 h. Given that JHA altered the m6A modification levels of BmSPI4 and BmSPI5, we therefore investigated whether JHA directly regulates the expression of BmSPI4 and BmSPI5 through m6A in BmN cells. According to the results of m6A sequencing, we identified that m6A peaks in the BmSPI4 CDS region ranged from NC_051385.1: 8,549,783-8,550,004, and the BmSPI5 CDS region ranged from NC_051385.1: 8,506,603-8,506,712 (Fig. 8C). We further generated reporter plasmids containing His-tag following the wild-type BmSPI4, BmSPI5, or mutant BmSPI4, BmSPI5 CDS. To avoid the influence of background expression of BmSPI4 and BmSPI5 in BmN cells, we designed specific qPCR primers located in the pIZ/V5-His vector, BmSPI4 and BmSPI5 CDS regions (Fig. 8D). We performed qPCR and western blotting to detect the expression of PizV5-BmSPI4 and PizV5-BmSPI5 with or without JHA treatment. The results suggested that JHA treatment significantly enhanced the expression levels of pIZ/V5-BmSPI4-WT and PizV5-BmSPI5-WT (Fig. 8E-F). However, JHA did not improve the expression of pIZ/V5-BmSPI4-Mut or pIZ/V5-BmSPI5-Mut (Fig. 8E-F). Taken together, the results revealed that JHA directly regulates BmSPI4 and BmSPI5 through m6A modification of CDS regions.
{kind=link}