Cellular expression of multiple Notch isoforms.
Previous work revealed the now canonical crystal structure of the mammalian Notch transcriptional complex containing fragments of RBPj, MAML1, and the N1ICD ankyrin domains in a head-to-head dimerized state12,13. Nam et al.12, also defined four key residues in the N1ICD ankyrin domain critical for dimerization of N1ICD molecules and determined that these residues are highly conserved in the NICD ankyrin domains of all four mammalian Notch isoforms. This data suggested that all Notch NICD domains likely engage in head-to-head dimerization interactions similar to N1ICD. Furthermore, given the overall sequence similarity of the Notch ankyrin domains and the conservation of these key amino acids required for dimerization (Supplementary Fig. 1), it was further hypothesized that NICD domains may also be able to engage in heterodimeric interactions, although this has not been investigated.
In order for heterodimeric NICD interactions to be even feasible, at least two isoforms of Notch receptors would need to be expressed in a single cell. To assess the expression of Notch receptors in single cell lines, and thus the possibility of NICD heterodimerization, we screened several cell lines for their potential for co-expression of Notch receptors. Reverse-Transcription PCR (RT-PCR) was used to monitor mRNA expression of each Notch protein. As shown in Fig. 1, each of the cell lines examined expressed mRNA for at least two Notch receptors, while several expressed three or even all four Notch receptors. These results suggested that cellular expression of multiple Notch isoforms is common and solidified the possibility that heterodimeric interactions between different NICDs is at least feasible.
Co-immunoprecipitation of NICD heterodimer complexes.
To test the hypothesis that the NICD domains can engage in heterodimeric interactions, we first established a cellular co-immunoprecipitation (Co-IP) strategy that involved the generation of FLAG, MYC, and HA-tagged versions of each of the four different NICD molecules. Co-transfection of 293T cells with two different tags would thus allow us to simultaneously monitor NICD homodimerization and heterodimerization in the precipitates. 293T cells were transiently co-transfected with cDNAs encoding one FLAG-tagged and a second MYC- or HA-tagged pair of NICD proteins. Anti-FLAG Co-IP was used to isolate FLAG-tagged NICDs, then immunoblotting with anti-MYC or anti-HA antibodies was used to detect MYC- or HA-tagged Co-IP NICD partners, depending on combination. As shown in Fig. 2A, full-length FLAG-N1ICD engaged in homodimeric interactions with full-length HA-tagged N1ICD and heterodimeric interactions with the HA-tagged N2, N3, and N4ICD molecules. Interestingly, N1ICD co-immunoprecipitated with N2, N3, and N4ICD better than with N1ICD itself suggesting that N1ICD preferentially engaged in heterodimeric interactions. Moreover, N1ICD most strongly precipitated with N4CID. N2ICD also formed homodimers and heterodimers with each of the other NICDs and again, the N2ICD-N4ICD interaction appeared to be the most robust (Fig. 2B). N3ICD formed heterodimers with each of the other NICDs (Fig. 2C) although dimerization with N1ICD and N2ICD appeared to be weak (Fig. 2C dark) and as before, the interaction between N3ICD and N4ICD appeared to be the strongest (Fig. 2C light). Finally, N4ICD demonstrated strong heterodimerization with the other NICDs, but again, N4ICD homodimerization appeared to be the most robust (Fig. 2D). Importantly, Co-IP of MYC and HA-tagged NICDs was consistently dependent on the presence of FLAG co-transfection thus illustrating the specificity of the co-immunoprecipitation system.
Collectively, this series of Co-IP experiments demonstrated that all NICDs appear to be capable of mix and match homodimer and heterodimer interactions. Moreover, these results also showed that the NICDs do not appear to equally engage in dimerization and that N4ICD consistently displayed the most robust homodimerization and heterodimerization compared to the other NICD molecules.
Ankyrin domains are not required for NICD Co-IP.
Although the canonical and transcriptionally active NICD tripartite complex is the most studied Notch dimer complex, another less well studied dimer complex was previously described by Vasquez et al.16, This alternative NICD dimer complex was identified by co-immunoprecipitation methods similar to those used above and was described as an “antiparallel” complex since it involved the dimerization to two N1ICD monomers via N-terminal to C-terminal interactions such that the dimerizing molecules were proposed to orientate in an antiparallel configuration reminiscent of a “yin-yang” symbol. In this yin-yang conformation, the ankyrin domains which mediate the tripartite complex head-to-head NICD interactions were found to be non-essential for dimerization16.
Based on these findings, it was important to determine if our co-immunoprecipitation experiments were detecting NICD dimers in the transcriptionally active head-to-head conformation, the yin-yang conformation, or perhaps a blend of both dimerization modalities. To accomplish this, we again performed the co-immunoprecipitation experiment with NICD molecules containing ankyrin domain mutations that are unable to form head-to-head dimer complexes. Arg1985 within the human N1ICD ankyrin domain was previously identified as important for NICD dimerization and transcriptional responses from promoters with paired RBPJ binding sites such as the Hes5 promoter12,13. Using sequence alignments, we found equivalent arginine residues in the mouse Notch1 and Notch4 NICDs (Arg1974 and Arg1685 respectively) (Supplementary Fig. 1) and performed site-directed mutagenesis to change these arginine residues to alanine residues. To confirm that these mutations decreased transcriptional activity from paired RBPJ sites and therefore head-to-head dimerization, we used a luciferase assay to monitor transcriptional activity from the SPS-containing Hes5 promoter. As shown in Fig. 3A, ankyrin mutant N1ICD demonstrated much weaker transcriptional activity compared to wild-type N1ICD. Moreover, ankyrin mutant N4ICD was also less efficient at driving Hes5 promoter activity, although the inherently low transcriptional activity of wild-type N4ICD made this effect less obvious compared to N1ICD. Since even non-dimerizing NICD domains minimally promote transcription, we were not surprised that these mutations failed to eliminate transcriptional activity altogether. Nonetheless, these results confirm that mutation of N1ICD and N4ICD at these positions decreases transcriptional activity by disrupting head-to-head dimerization of NICDs.
We next sought to determine if ankyrin domains are required for NICD co-immunoprecipitation. To accomplish this, we again performed the Co-IP assay to compare the ability of wild-type and ankyrin mutant N1ICD and N4ICD to Co-IP in 293T cells (Fig. 3B). 293T cells were transiently co-transfected with cDNAs encoding one FLAG-tagged and a second MYC- or HA-tagged pair of NICD molecules. Anti-FLAG Co-IP was again used to isolate FLAG-tagged NICDs, then immunoblotting with anti-MYC or anti-HA antibodies was used to detect MYC- or HA-tagged Co-IP NICD partners, depending on combination. Due to inconsistencies in observing full-length N1ICD homodimers on Western Blots, which we will discuss later, we found that a C-terminally truncated N1ICD construct lacking its PEST domain more reliably precipitated was was employed for this experiment. Despite the importance of ankyrin domains for transcriptional activity and dimerization, we were unable to detect any change in N1ICD or N4ICD homodimerization when the ankyrin domains were mutated. This result suggested that ankyrin domains are not required for co-immunoprecipitation but did not rule out the possibility that ankyrin domains may at least participate in Co-IP. To determine if ankyrin domains are capable of mediating Co-IP, we therefore compared the ability of HA-tagged N4ICD to Co-IP with various isolated fragments of the N1ICD. Consistent with the results of Vasquez et al.16, we found that full-length N4ICD was able to interact with an N-terminal fragment of N1ICD containing just the RAM/Ank domains as well as with the isolated N1ICD ankyrin domain itself (Fig. 3C). Furthermore, as shown in Fig. 3D, complete ablation of the N4ICD ankyrin domain reduced, but did not abolish Co-IP between N4ICD molecules. Taken together, these results suggested that NICD ankyrin domains are not required for Co-IP between NICD complexes but are capable of mediating Co-IP and therefore, that our Co-IP procedure was likely recovering both the head-to-head and yin-yang dimer complexes.
NICD molecules heterodimerize on DNA.
Our results indicated that dimerization between NICD molecules (as measured by Co-IP) did not directly correlate to transcriptional activity, that ankyrin domains were not required for Co-IP, and yet that isolated ankyrin domains could dimerize. Taken as a whole, these results suggested that NICDs may simultaneously exist in multiple dimerization states. One state was detected by co-immunoprecipitation but did not apparently correspond to transcriptional activity (yin-yang dimers) while the second state appeared to be transcriptionally active and dependent on interactions between ankyrin domains (head-to-head dimers). These observations raised the important question whether or not any of the NICD complexes that were being recovered by Co-IP were engaged in heterodimer complexes on DNA or rather were strictly in a non-transcriptional yin-yang complex as suggested by Vasquez et al16. To address this question, we adapted our Co-IP procedure into a chromatin IP experiment to determine if any co-immunoprecipitating NICDs were bound to DNA. 293T cells were co-transfected with combinations of FLAG-tagged N1ICD or N4ICD coupled with HA-tagged N1ICD or N4ICD and simultaneously with a plasmid containing either the Hes4 or Hes1 SPS promoters. After crosslinking and DNA fragmentation, cell lysates were immunoprecipitated first with anti-FLAG antibodies then with anti-HA antibodies such that DNA would only be recovered if it was associated with both FLAG and HA-tagged NICD proteins. Control samples included transfection with FLAG-tagged N1ICD alone and a single anti-FLAG immunoprecipitation step (positive control) and a negative control that was transfected with HA-tagged NICD and subjected to the two-step Co-IP procedure. In all cases, precipitated DNA was detected by PCR with oligos that were designed to flank the paired head-to-head SPS binding sites within the Hes4 and Hes1 promoters. As shown in Fig. 4A, both Hes1 and Hes4 sequences were detected in the precipitated chromatin of all NICD combinations and the ChIP was specific since Hes4 and Hes1 promoters were not recovered if a FLAG-tagged protein was omitted.
On the surface, this result seemed to confirm that NICD molecules engage in homodimer complexes but also supported the hypothesis that NICD heterodimers may form on DNA. Despite this, it was important to investigate whether the captured NICD molecules were actually engaged in dimerized complexes or instead were individually binding to DNA in a non-dimerized state. To investigate this possibility, we performed another Co-IP experiment comparing NICD binding to SPS promoters with 16 or 21 bp (base pair) gaps separating the RBPj binding sites. 16 bp gaps have been shown to be optimal for supporting dimerization and transcriptional activity whereas 21 bp gaps are too wide to support dimerization and only very weakly promote transcription17. In addition, we also compared wild-type NICD proteins to their transcriptionally inactive ankyrin mutant versions. As shown in Fig. 4B, NICD homodimer and heterodimer complexes were detected on both the 16 and 21 bp gapped SPS sites. Moreover, the recovery of DNA in the ChIP assay was also independent of transcriptionally inactivating mutations in the ankyrin domains. These results indicate that heterodimer complexes do exist on DNA, but since the recovery of dimer complexes was independent of both gap length and ankyrin domain function, this ChIP analysis was unable to determine if heterodimer NICD pairs are transcriptionally active.
Head-to-head heterodimerization of NICD complexes affect transcriptional activation.
Thus far, our results demonstrated that NICDs can engage in heterodimeric complexes in both the non-transcriptionally active yin-yang conformation as well as in the transcriptionally active head-to-head dimer conformation. However, none of our experiments had been able to determine if the head-to-head heterodimer complexes that form on DNA were in fact transcriptionally active. Assessing the transcriptional activity of heterodimer complexes is somewhat challenging since a simple reporter assay cannot distinguish between the transcriptional activities of homodimer and/or heterodimer complexes. Thus, we required a system where we would be able to directly monitor the transcriptional activity of heterodimers while excluding the transcriptional activity of homodimers. Drawing inspiration from Liu et al.15, and Nam et al.12, we created a series of compensatory NICD mutants that were incapable of homodimerization but still capable of heterodimerization. This strategy is based on the fact that NICD dimerization is mediated in part by salt bridges between positively charged lysine (K) residues and negatively charged glutamic acid (E) residues that interact across the interface of adjoining ankyrin domains as schematically illustrated in Fig. 5A and 5B. Nam et al previously demonstrated that K1946E (EE) and E1950K (KK) mutation of human N1ICD thwarted homodimerization of EE or KK species but permitted homodimerization between N1ICD EE and KK species through restored salt bridge interactions across the interface between adjacent ankyrin domains. We hypothesized that a similar approach would allow us to monitor heterodimer formation between different species of mouse NICD molecules. To accomplish this, we first aligned the human N1ICD ankyrin domain with the ankyrin domains of the mouse Notch proteins to find the corresponding residues (Supplementary Fig. 1). Site directed mutagenesis was then used to mutate the mouse NICD molecules to contain lysine only (KK) or glutamic acid only (EE) residues at these dimerization sites. Specific mutation sites in each NICD are listed in Fig. 5C.
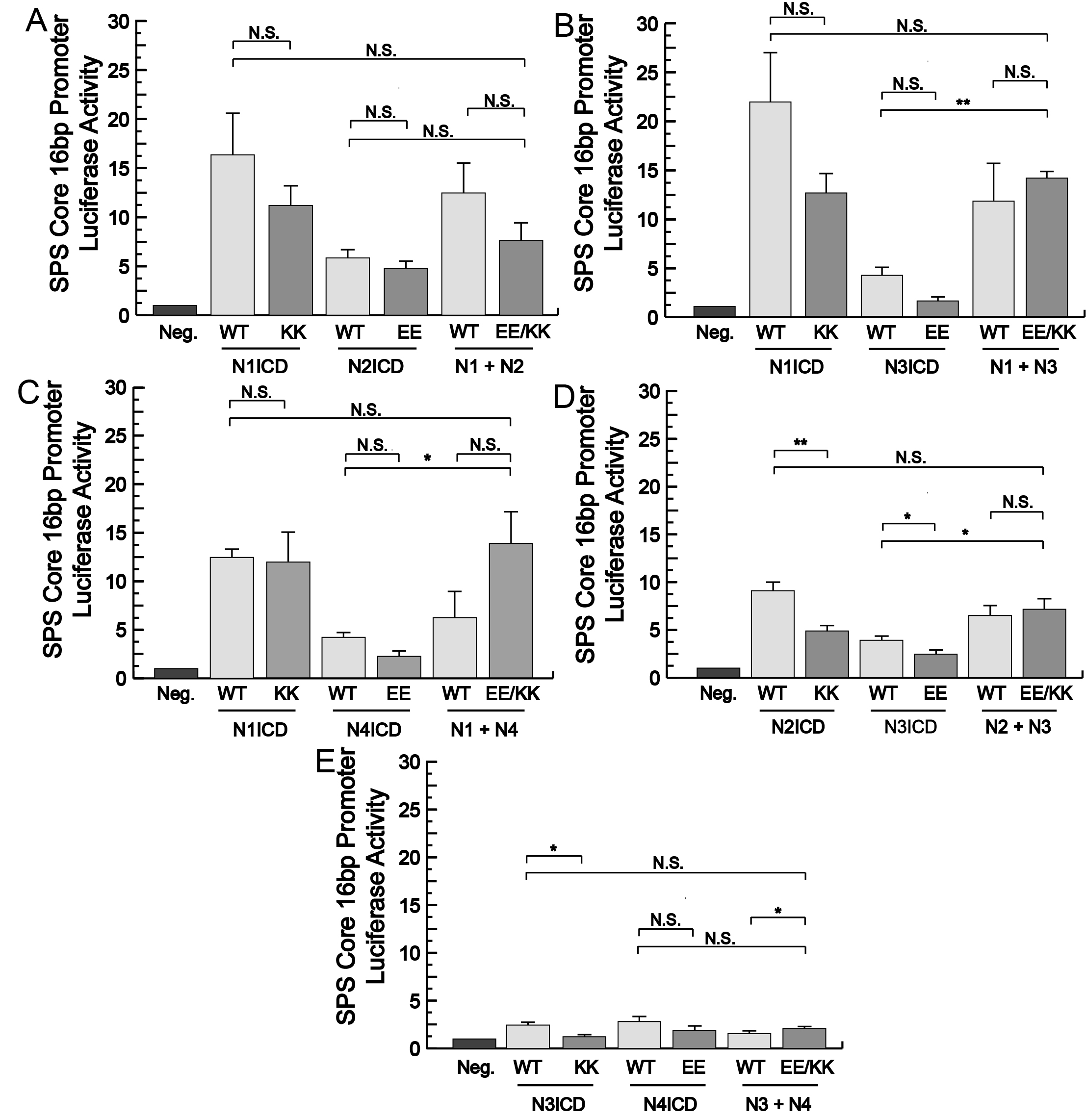
To test these mutants, we co-transfected 293T cells with various combinations of EE and KK mutants and luciferase reporters that have 16 bp gaps between their RBPj binding sites. The 16 bp gap has been shown to be the optimal distance between dimerizing NICDs to enable synergistic and robust transcriptional activity17. As shown in Fig. 6, the single N1ICD KK mutant we managed to generate did not significantly reduce activity compared to the wildtype N1ICD and for unknown reasons, we were never able to generate the second EE mutant in N1ICD. Despite this, the EE and KK mutants of both N2ICD and N3ICD significantly reduced luciferase activity. Finally, the EE but not KK N4ICD mutant, also considerably diminished reporter activity compared to control N4ICD.
Having found NICD mutations that were unable to support NICD homodimerization, we next sought to determine if these same mutant NICDs would be able to rescue transcriptional activity when KK and EE versions were combined. To accomplish this, we again co-transfected 293T cells with the 16 bp gap luciferase reporter and various EE and KK combinations of mutant NICDs. We found that some but not all mutant NICD combinations were able to support transcription. For example, N2ICD KK and N4ICD EE both have reduced transcriptional activity compared to their non-mutated NICD counterparts indicating their inability to form homodimers, but together this NICD pair supported transcriptional activity comparable to wild-type N2ICD homodimers alone (Fig. 7A). A similar result was also observed when N2ICD KK and N3ICD EE mutants were co-transfected (Supplementary Fig. 2). Importantly, The N2ICD and N4ICD pair was unable to support strong transcriptional activity on a promoter with 21 bp between its RBPj binding sites (Fig. 7B). The longer base pair gap between TP-1 elements does not support synergistic and robust transcriptional activity, therefore this result indicated that the NICDs were not working individually to activate robust transcription. Despite the success of the N2ICD KK and N4ICD EE dimer pair and the N2ICD KK and N3ICD EE dimer pair, other KK/EE combinations failed to rescue robust transcriptional activity (Supplementary Fig. 2). Nonetheless, this data strongly supports the conclusion that the aforementioned NICD combinations were forming legitimate heterodimers that synergistically activated transcription in a manner similar to normal homodimer pairs.
Notch dimerization affects promoter activity with varying specificity.
KK and EE compensatory mutations in the dimer domains of N2ICD and N4ICD rescued transcriptional activity using the optimized 16 bp SPS promoter suggesting heterodimerization between N2ICD-KK and N4ICD-EE. This promoter, however, is an artificially generated and optimized SPS element17 that does not contain flanking DNA sequences. Therefore, it was important to determine if these NICD pairs were also able to heterodimerize on more natural Notch-specific promoters. Using the same N2ICD-KK and N4ICD-EE mutants, we performed luciferase assays on the Hes1, Hes4, and Hes5 promoters that not only contain the SPS element, but also hundreds of base-pairs of flanking DNA sequences. Consistent with previous results17, the Hes1 and Hes4 promoters were only weakly activated by N2ICD or N4ICD transfection and interestingly, the EE and KK mutants had transcriptional activities comparable to their wild-type counterparts suggesting that these mutations failed to block homodimerization on these promoters. On the other hand, the N2ICD-KK but not the N4ICD-EE mutant was unable to form homodimers on the Hes5 promoter (Fig. 8). Surprisingly, the combination of the N2ICD-KK and N4ICD-EE mutants that rescued transcriptional activity of the synthetic promoter failed to rescue transcriptional activity on the Hes5 promoter.
{kind=link}