Beyond the association between obesity and cardiovascular risk, current knowledge supports a link between obesity and incidence of cancer and poorer survival outcomes compared to normal (healthy) weight subjects (20). This evidence highlights the possible impact of obesity and particularly the putative effect of hypertrophied adipocytes in the HL TME.
Distinctive environmental factors, particularly the excess BM adiposity, may influence absolute and relative cellular population distribution at the marrow microenvironment (5). This unbalanced microenvironment might support recruitment of malignant cells, promoting BM metastasis formation, and maintenance of a specific immunosuppressive environment that supports tumor growth (5, 21, 22). As seen in HL, the BM is one of the common sites for metastasis (24). However, the knowledge about the profile of adipokines in the BM fluid from subjects with cancer and/or with obesity remains limited. Therefore, considering the obesity context, we assessed the adipokine profile as a gauge of local adipose tissue activity.
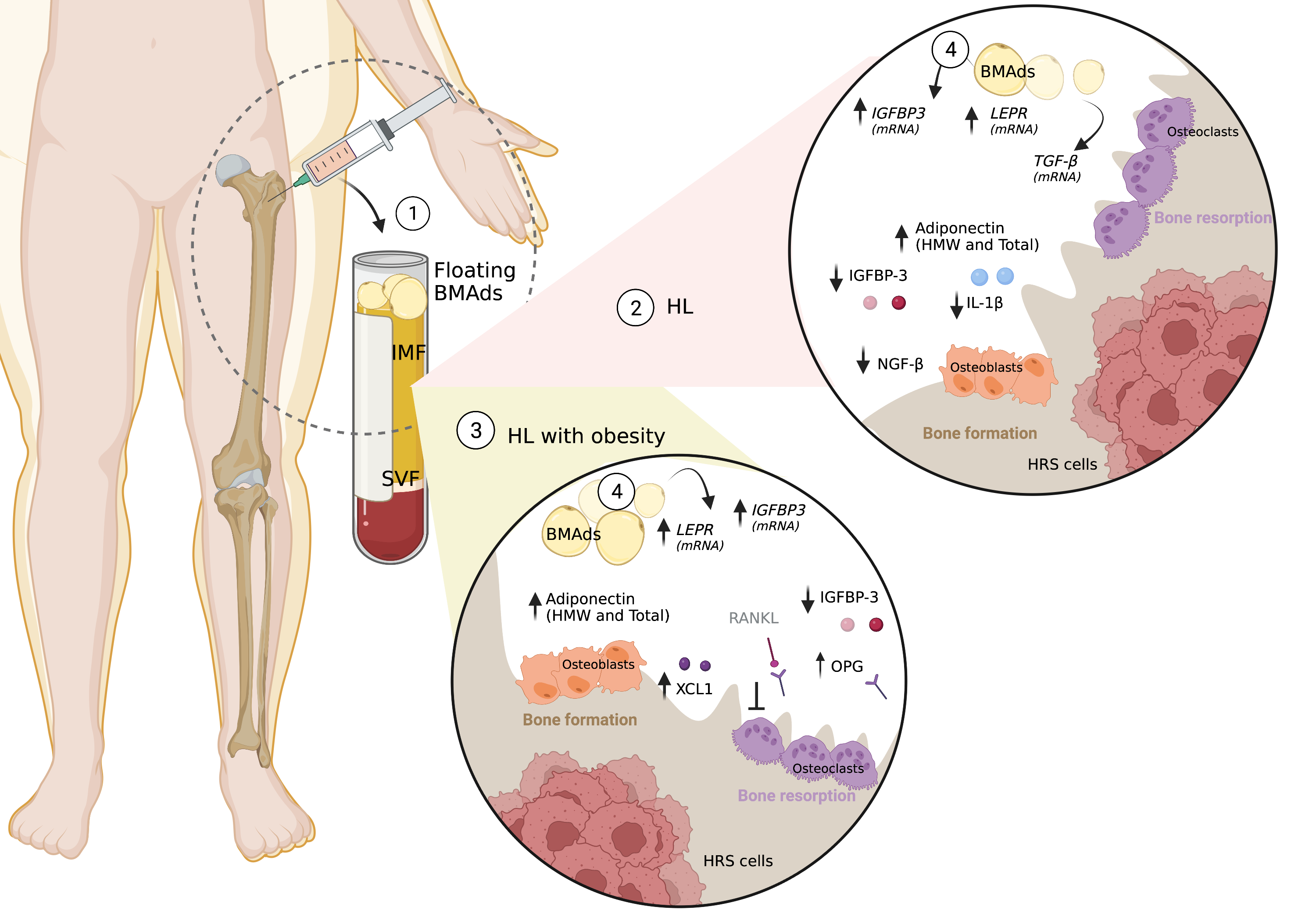
By comparing adipokine’s expression in cancer and / or adiposity status, our findings revealed that adipokine expression from several axes were downregulated in the IMF of HL subjects, namely cytokines (IL-1β, IL-1α, IL-6sR, IL-8, IL-12, Fas/CD95, MIP-1b/CCL4), sTNFRII, chemokine ligands (CCL-2 and CCL-3, CCL-16), IGF (IGFBP-1, IGFBP-2, IGFBP-3, IGF-1sR), leptin, growth factors (TGF-β, FGF-6) and bone remodeling molecules (OPG). Lower levels of pro-inflammatory cytokines were found in the IMF of HL patients, supporting the presence of an immunosuppressive microenvironment. IL-6sR, IL-12 and sTNFRII are associated with inflammatory processes at the BM HL niche and have been detected in other hematological malignancies, such as myeloproliferative neoplasms (25). IL-6 has been implicated in oncogenesis and metastasis, throughout osteoclast differentiation and consequent release of growth factors, as observed in multiple myeloma (26). IL-12 has been implicated as stimulator of diseases characterized by inflammation-induced bone destruction. This cytokine stimulates interferon-γ (IFN-γ) synthesis through Signal Transducer And Activator Of Transcription 4 (STAT4) activation promoting Th1 helper cells differentiation with important role on cancer treatment, by controlling tumor growth (27). Indeed, treatment with IL-12 transduced BM cells has been shown to impact metastatic prostate cancer (28). Moreover, murine second generation CAR T cells expressing IL-12 were capable of eradicating established B cell lymphoma with a long-term survival rate of ∼25% (29), supporting an anti-tumor effect of this cytokine. Findings suggest that decreased expression of IL-12 at HL BM TME might represent an immune escape mechanism. Another immune escape process involves the expression of Fas (CD95) by cancer cells, which modulates apoptosis through binding at T cells ligand Fas-ligand (FasL), inducing T cell death. Here, we observed decreased Fas expression in HL, independently of obesity status, which may be associates with anti-apoptotic signal driven by CD95+ cytotoxic T cells (30).
It is known that immune escape mechanisms of HRS cells are notable, protecting them by inhibition of cytotoxic T and NK cells activity, as well as promoting anti-inflammatory Treg and Th2 cells recruitment and differentiation (31, 32). Therefore, it is acceptable that at the bone marrow niche we found a counter-regulatory mechanism favoring a protective microenvironment for malignant HRS cell seeding, as an immune escape mechanism.
The IGF axis has also been implicated in obesity-cancer association (33). While IGF-1 is produced at the tumor microenvironment and HRS cells express IGF-1R, the ligand also binds to specific proteins to form binary complexes with binding proteins (34, 35). Here, we found a significantly reduced expression of IGBPs − 1, -2 and − 3 at the medullar fluid of HL subjects compared to donors. De-regulation of IGFBPs has also been demonstrated in the BM of multiple myeloma (36). In particular, the IGFBP-3 protein was consistently reduced in the IMF of HL patients, independently of the obesity status, which might result in an increased free IGF-1 levels and availability of this pro-proliferative ligand. To understand the role of IGFBP-3 in the BM TME of HL subjects, we conducted in vitro studies using human recombinant IGFBP-3 and IGF-1 proteins. Previous work on L428 cells demonstrated that IGF-1 [16 ng/mL] and [40 ng/mL] induced cancer cell growth (35). However, for IGFBP-3 there is not much evidence in HL cell lines, which prompted us to test the concentration of IGFBP-3 that we found in IMF from HL patients and reported concentrations from healthy subjects (37). We observed a positive proliferative effect of IGF-1 in HRS cells, but this effect was no longer observed when IGFBP-3 was added, suggesting a competitive inhibition effect. Furthermore, we observed that the combination of IGF-1 [16 ng/mL] and/or IGFBP-3 [15 ng/mL] decreased the impact of this insulin growth factor binding protein in apoptosis. In other cancer types, the IGFBP-3 regulated IGF-1 by blocking IGF-1R with implications on transcription level of type 1 insulin-like growth factor receptor (IGF1R), ERK and AKT (38, 39). Furthermore, at protein level, the interaction between IGFBP-3 and IGF-1 decreased the activation of ERK and AKT, although individually, IGFP-3 induced the activation of mTOR and IGF-1 AMPK. It is described that IGFBP-3 interacts with its receptor (IGFBP-3R/TMEM219) inducing apoptosis (40). However, the observed data may be dependent on availability of each molecule or the pleiotropic nature of IGFBP-3, as proposed elsewhere (40).
Interestingly, although lower levels of IGFBP-3 were observed at BM fluid, its expression may be modulated by BMAds, since we observed a higher mRNA transcript of IGBP-3 in HL patients in relation to donors, excluding in obesity groups (HL OW/OB versus Donors OW/OB). It has been previously described that IGFBP3 expression is regulated during adipocyte differentiation by interfering with the PPAR gamma-dependent processes (41). Remarkably, an increased expression of IGFBP-3 in the BM has been reported to facilitate bone metastasis by increasing TGF-β-mediated cell proliferation (41). Moreover, the IGFBP-3-modified BM-derived mesenchymal stem cells, promoting apoptosis and downregulating the expression of B-cell lymphoma-2 (Bcl-2), but increasing the expression of the pro-survival Bcl-2 associated X protein in human pulmonary artery smooth muscle cells (42). From our cohort, the increase expression of IGFBP-3 and TGF-β in adipocytes, suggest that they contribute to the formation of an immunosuppressive microenvironment, although we cannot infer an association to bone marrow invasion.
Bone resorption and bone formation are likely to become affected (23). Concerning obesity, and when patients were stratified by BMI classification, we observed osteoprotegerin (OPG) was significantly elevated in the IMF of HL OW/OB compared with lean HLs. OPG is a secreted glycoprotein member of TNF-alpha receptor superfamily, which acts in bone microenvironment as a decoy receptor, binding to RANKL-mediated osteoclast recruitment and activation. Therefore, the increased RANKL/OPG ratio is critical for osteoclastogenesis, by inhibiting osteoclasts differentiation and osteoclastic bone resorption (43). Our findings may reflect a systemic influence of the environment either modulated systemically, by the tumor itself, or through paracrine and/or endocrine mediators from adipocytes and adipose tissue. It is well established that OPG is secreted primarily by osteoblasts but also by adipocytes (44). In conditions of excess adiposity, some reports mentioned increased BMAd count and hypertrophy (45). Adipocytes, normal constituents of the BM microenvironment, were reported to crosstalk with osteoblasts and increase RANKL, while decreasing OPG secretion in the BM, influencing RANKL/OPG ratio through a still unknown mechanism (46). Previous data supports a reduced level of OPG and increased RANKL in the bone marrow of obese mice, which were correlated with higher BM adiposity and upregulated osteoclastogenesis (47, 48). In agreement, previous work revealed higher OPG levels in the IMF of osteoporotic and leaner women (49). Correlating with other cytokines that we found relevant at IMF, the downregulation of pro-inflammatory cytokine IL-11 and VEGF-A may also contribute to inhibition of bone resorption. IL-11R has been suggested as a possible cell surface target for ligand-directed applications in human leukemia and lymphoma (50). On the other hand, VEGF serves as a survival factor for chondrocytes and couples the resorption of cartilage with bone formation during endochondral ossification. Recently, VEGF has also been found to regulate the balance between osteoblast and adipocyte differentiation in BM mesenchymal stem cells (51).
Beyond the adipo-like molecules described previously, adiponectin is highly enriched at BM microenvironment. Although BM adipose tissue is a source of adiponectin, circulating adiponectin decreases in obese, and insulin-resistant states (52). Since adiponectin exists in distinct multimeric forms including low-molecular-weight (LMW) trimers, middle-molecular-weight (MMW) hexamers, and high-molecular-weight (HMW) complexes, we quantified these isoforms at IMF. We observed higher levels for HL patients with obesity, comparing to donors, which confirms the unfavorable niche for metastasis since low adiponectin levels represents a risk factor for cancer, and for a more aggressive phenotype (53, 54). Furthermore, BMAds of HL patients do express significantly higher levels of adiponectin mRNA. BMAds might be particularly susceptible to such stress, given their relatively high expression of proinflammatory genes and relative resistance to insulin (53). Thus, it seems likely that obesity would also lead to adipocyte dysfunction within the BM, thereby compromising production of adiponectin (55).
Although, we didn´t found a metastatic niche in the studied patients, we found increased serum levels of HGF and MMP-9, precursors of invasion and of metastasis. Evidence reported that the MMP-9 expression by neoplastic cells in HL is associated with EBV positivity (56). This association is correlated with increased serum levels of MMP-9 in HL patients (57). In our study, we also found increased serum levels of MMP-9, a matrix metalloproteinase regulator, involved on degradation of extracellular matrix, growth factors activation and their expression are associated with immune system regulator (58). Notably, treatment with anti-MMP-9 and anti-PDL1 antibodies reduced T-cell receptor clonality and increased TCR diversity (58). Nevertheless, other factors associated with invasion and metastasis, activation of stemness, clonal expansion, and cell transformation, such as HGF and MMP9, were increased in HL patients’ serum comparing to controls. This profile seemed to be associated to particularities of the disease, since we found the same profile when considering the normal ponderal subjects, comparing HL patients with controls. Furthermore, the decrease of IL-8, a chemotactic signal for neutrophils, may also indicate a non-reactive infiltrate in HL disease. In line with this, the downregulation of NGF-beta in BM of HL patients seemed to corroborate the decrease differentiation of BM mesenchymal stem cells, thereby diminishing the capability of metastasis (59). The “seed and soil hypothesis” explains the mechanism by which a certain primary cancer metastasis to a specific organ, and according to this theory (60), in Hodgkin Lymphoma disease we might not have good “soil” (bone), for “seed” (cancer cells).
{kind=link}