Bacterial enumeration
During dried shrimp production, as the shrimp dehydrated, their volume gradually shrank (Fig. 1A). The pH level significantly decreased at ME1 stage and slightly increased thereafter (Fig. 1B). TVC significantly increased at ME3 stage (Fig. 1C).
Diversity of the bacterial community in dried shrimp
An average of 79,985 paired-end reads per sample were obtained, and the numbers of taxon tags ranged from 45,685 to 134,259 among the samples with 73,329 taxon tags per sample (Table S1). These tags were grouped into 270 microbial Operational Taxonomic Units (OTUs) (Table S1).
The number of OTUs in ME0 period samples was the highest, and a total of 170 OTUs was shared by all three periods (Fig. 2A). The Richness (Fig. 2B) and Shannon (Fig. 2C) indices indicated higher α-diversity in the ME0 period than in the latter two periods. PCoA analysis also suggested that samples within groups tended to cluster together, while the ME0 period was distinct from the latter two stages (Fig. 2D). During dried shrimp production, the similarity between samples within groups gradually decreased, indicating a trend towards unstable community structures, and significant differences were observed in the community structures among different sample groups (Fig. 2E). A previous study demonstrated the importance of the initial microbial load found on ready-to-eat foods. However, it should be noted that the microbiological load of these foods at the point of sale can be influenced by several factors, including processing, storage, and display conditions (Angelidis et al., 2006; Beuchat and Ryu, 1997).
Microbial community composition and changes during preparation
In the ME0 period, the predominant bacterial phyla in the samples were Bacteroidetes, Actinobacteria, and Proteobacteria, while in the latter two periods, Proteobacteria was the predominant phylum, and the relative abundance of Firmicutes was increased (Fig. 3A). At the genus level, the ME0 samples showed a high abundance of Psychrobacter, while the ME1 samples mainly included Vibrio and Photobacterium genera, and the genera Vibrio, Shewanella, Lactococcus, and Photobacterium were more abundant in the ME3 samples (Fig. 3B). A previous study showed that Firmicutes and Proteobacteria were the predominant phyla during the drying process in dry sausages, consistent with our results (Hu et al., 2020). Lactobacillus sakei is the predominant species in Croatian dry fermented sausages (Zgomba Maksimovic et al., 2018).
Indicator species analysis indicated that the abundance of Rhodobacter, Flavobacterium, Pseudoalteromonas, Psychrobacter, and Loktanella genera was significantly higher in the ME0 period than in the latter two stages, while Aliivibrio and Vibrio were more abundant in the latter two periods (Fig. 3C). Photobacterium was significantly more abundant in the ME1 period than in the other two stages, whereas Shewanella was significantly less abundant in the ME1 period than in the other two periods (Fig. 3C). In particular, Lactococcus gradually increased during shrimp drying process (Fig. 3C). Vibrio, Photobacterium, and Shewanella are known marine animal pathogens and potential opportunistic human pathogens, whereas Lactococcus are potential beneficial bacteria. A previous study showed that Aliivibrio and Vibrio are important pathogens among seafood and pose a threat to human health (Huehn et al., 2014; Neetoo et al., 2022; Toranzo et al., 2023). Another study showed that Photobacterium and Shewanella were the most prevalent and abundant pathogens among bluespotted seabreams, making up 30.2% and 11.3% of detected pathogens (Itay et al., 2022). Lactococcus is the most important beneficial bacteria in dried seafood. One study found that Lactococcus lactis 69, isolated from sun-dried meat, could secrete a heat-stable bacteriocin that exhibited good inhibitory effects againast various pathogens (Biscola et al., 2013). Another Lactococcus lactis KTH0-1S isolated from dried Thai shrimp could produce a heat-stable bacteriocin that also inhibits food-borne pathogens and reduces tyramine accumulation (Saelao et al., 2018, 2017). The contribution of Lactococcus to flavor development in fermented meat products was found to be relatively insignificant, primarily owing to its low lipolytic activity (Flores and Toldrá, 2011). By contrast, the accumulation of Lactococcus during the drying process in shrimp plays an important role in changes in the shrimp microbiome and flavor.
Potential functions of the bacterial community
Predicted functions results showed that the aerobic category was significantly higher in the ME0 period than in the latter two periods (Fig. 4A), and the aerobic category decreased during the shrimp drying process. Facultatively anaerobic (Fig. 4B), potentially pathogenic (Fig. 4C), and stress tolerant (Fig. 4D) categories were significantly higher in the latter two periods, while anaerobic (Fig. 4E) and biofilm formation (Fig. 4F) categories did not show significant differences among the three periods. One study showed that aerobic microorganisms such as E. coli O157:H7 are reduced by three logarithms during the process of drying apple slices (Derrickson-Tharrington et al., 2005). Another study proved that potential pathogens undergo osmotic stress-tolerances increases to adapt to environmental changes during drying in the food chain (Burgess et al., 2016; Sleator and Hill, 2002).
Metabolomic changes in dried shrimp during preparation
A total of 13,713 metabolites were identified in the samples during shrimp drying, with 7,571 metabolites identified at the secondary level (Table S2). Among the differentially accumulated metabolites (DAM), the majority increased during shrimp drying (Fig. 5A). Pairwise comparison of differentially regulated metabolites was performed across the three periods, and 717 metabolites were found to be differentially regulated (Fig. 5B). Compared to the ME1 period, the arachidonic acid metabolism, purine metabolism, the sphingolipid signaling pathway, biosynthesis of nucleotide sugars, drug metabolism and other enzymes, the apelin signaling pathway, the pentose phosphate pathway, nicotinate and nicotinamide metabolism, arginine and proline metabolism, the cAMP signaling pathway, and calcium signaling pathway were significantly upregulated in ME0 samples (Fig. 5C). Compared with the ME1 period, the biosynthesis of unsaturated fatty acids, arachidonic acid metabolism, lysine degradation, necroptosis, phenylalanine metabolism, lysine biosynthesis, fatty acid elongation, propanoate metabolism, glycerophospholipid metabolism, linoleic acid metabolism, valine, leucine and isoleucine biosynthesis, sulfur metabolism, aminoacyl-tRNA biosynthesis, and the prolactin signaling pathways were significantly upregulated in ME3 samples (Fig. 5D). Biosynthesis of unsaturated fatty acids is an important change during the sun-drying of shrimp. A previous study showed that unsaturated fatty acids (including monounsaturated fatty acids and polyunsaturated fatty acids) were released by Saccharomyces cerevisiae 31 during the fermentation process of fish paste (Chen et al., 2017). One study found 43.90% monounsaturated fatty acids, 28.61% polyunsaturated fatty acids, and 27.48% saturated fatty acids among dried shrimps (Sampaio et al., 2006). The arachidonic cis-5, 8, 11, 14-Eicosatetraenoic (C20:4 n-6) value in the raw shrimps was increased significantly by smoking and sun-drying (Akintola, 2015). Amino acid metabolism also plays a vital role in the drying of shrimp. A previous study showed that tyrosine values increased approximately 70% and 80% respectively in smoked and sun-dried shrimp, which was associated with phenylalanine metabolism and lysine biosynthesis (Akintola, 2015; Omolola, 2013).
Temporal analysis of DAM was performed by STEM, which identified three significant profiles with an overall increasing trend (Fig. 6A). The DAM in these significant profiles were mainly enriched in oxidative phosphorylation, cGMP-PKG signaling, flavones and flavonols biosynthesis, biosynthesis of various antibiotics, novobiocin biosynthesis, purine metabolism, AMPK signaling, Type I polyketide structures, riboflavin metabolism, flavonoid biosynthesis, chlorocyclohexane and chlorobenzene degradation, nicotinate and nicotinamide metabolism, and longevity regulating pathway, etc (Fig. 6B). Flavones and flavonols are important nutriments impacted by food among the fermentation and drying proceses. A previous study showed that the total flavonoid content of green tea was highest after drying at 38 ℃ (Roshanak et al., 2016).
Interaction of key microbiota and metabolites in dried shrimp
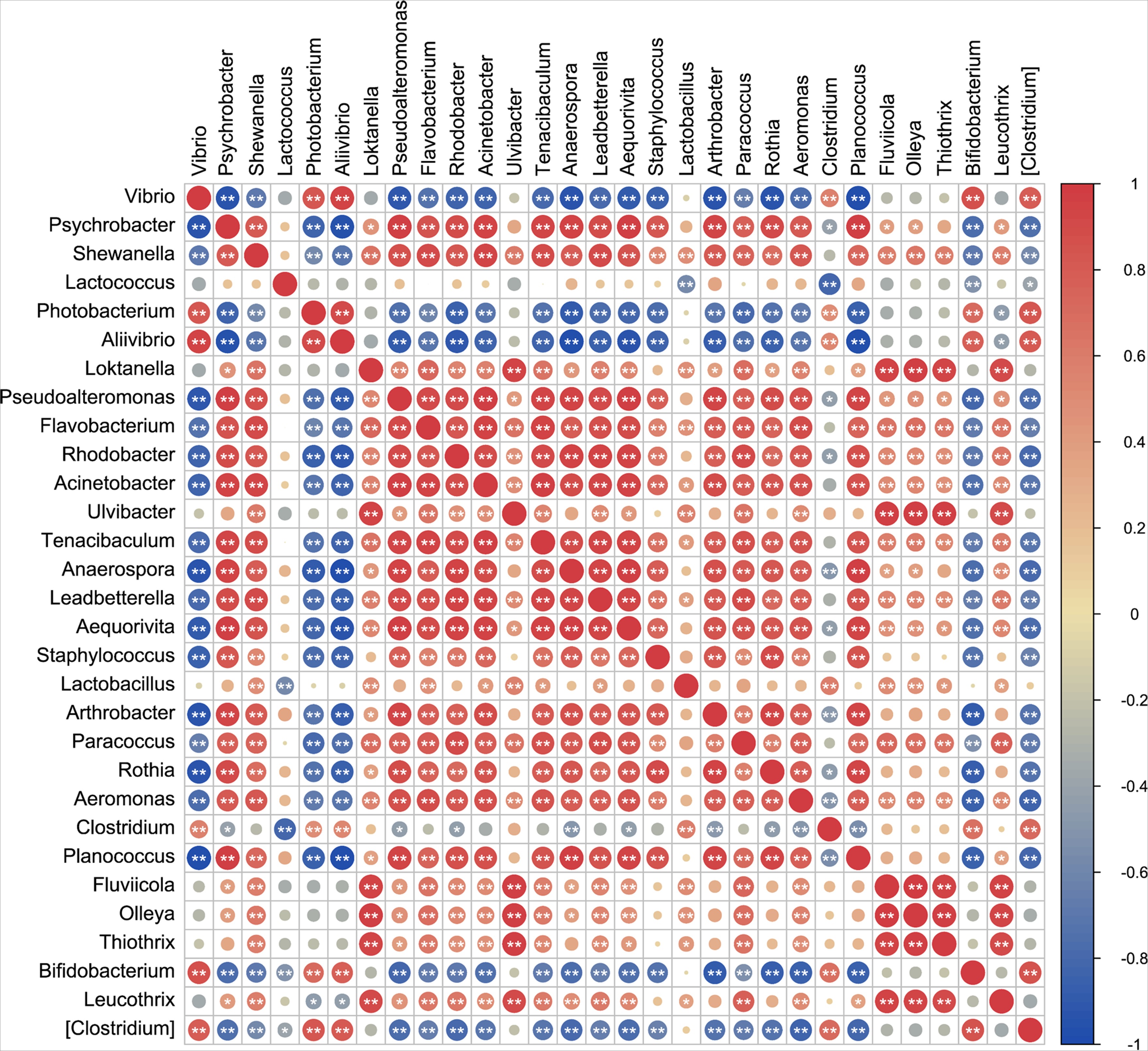
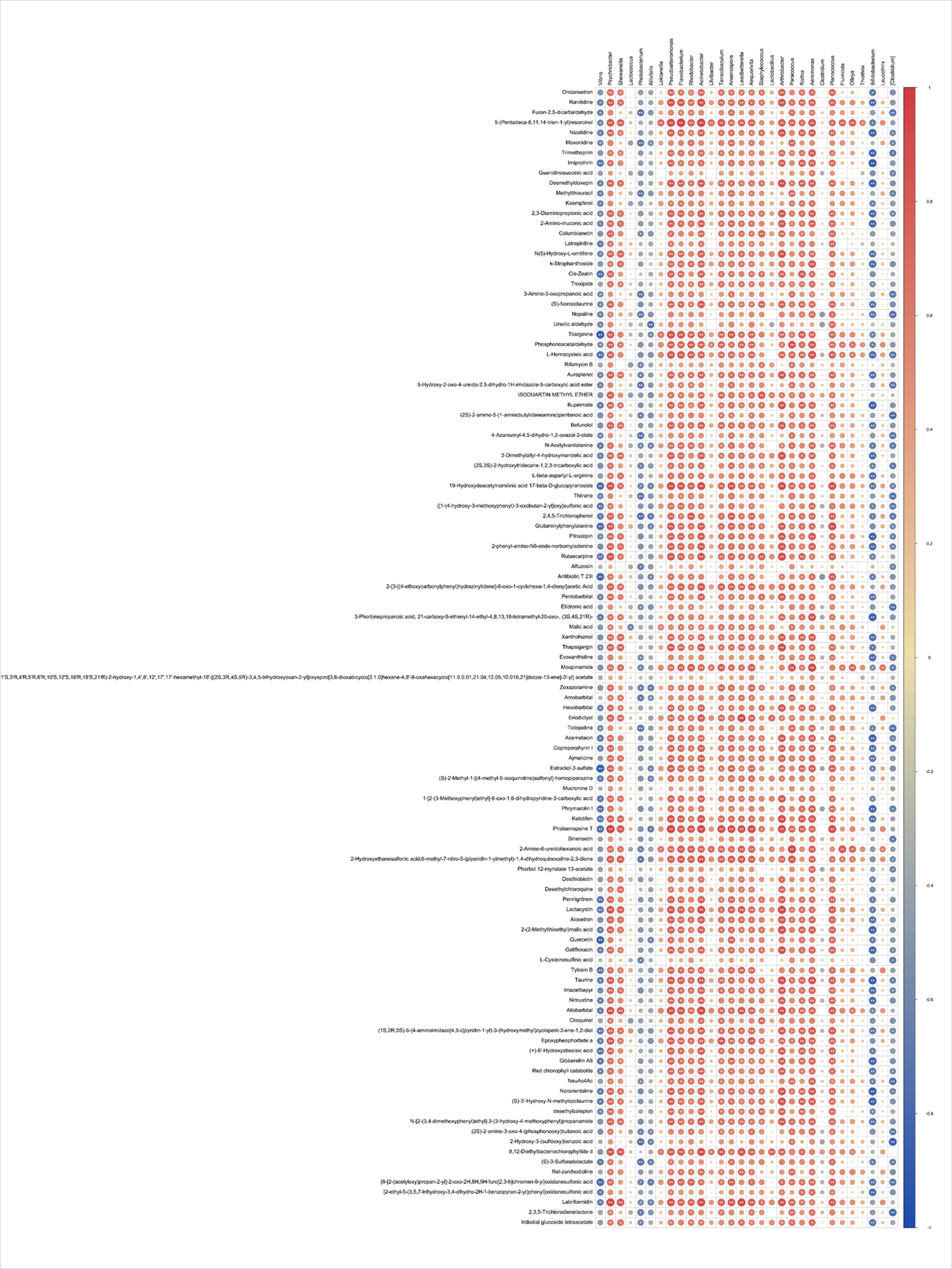
Spearman’s correlation analysis was performed on the key indicator microbial community and on differential metabolites (Figures S1 and S2). The majority of key microbial taxa, including Psychrobacter, Shewanella, Lactococcus, Flavobacterium, Rhodobacter, Acinetobacter, Ulvibacter, Tenacibaculum, Leadbetterella, Aequorivita, Staphylococcus, Lactobacillus, Arthrobacter, Paracoccus, Aeromonas, Fluviicola, Olleya, Thiothrix, and Leucothrix, showed positive correlations with each other, and exhibited positive correlations with differentially accumulated metabolites (Figures S1 and S2). By contrast, Vibrio, Allivibrio, Photobacterium, and Bifidobacterium were positively correlated with each other, but their abundances were predominantly negatively correlated with differentially accumulated metabolites (Figures S1 and S2). Based on the results of these correlation analysis, a network depicting microbial interactions and microbial-metabolite interactions was constructed (Fig. 7). The results indicate that Bifidobacterium, Clostridium, and Photobacterium are mainly associated with heterocyclic compounds and organic acids and their derivatives metabolites in a co-occurring manner, while Arthrobacter and Staphylococcus were predominantly mutually exclusive with heterocyclic compounds (Fig. 7). Previous studies have shown that Bifidobacterium and Photobacterium are associated with heterocyclic compounds (Faridnia et al., 2010; Xu et al., 2000). Meanwhile, the immobilization of Clostridium was the main effect of organic acids (Dolejš et al., 2014; Stinson and Naftulin, 1991). One study showed that Arthrobacter mainly degrades heterocyclic compounds (Guo et al., 2019). Another study showed that Staphylococcus is primarily exclusive to heterocyclic compounds (Paudel et al., 2017).
{kind=link}
{kind=link}