CHI3L1 is highly overexpressed in glioblastoma and is associated with the mesenchymal subtype markers
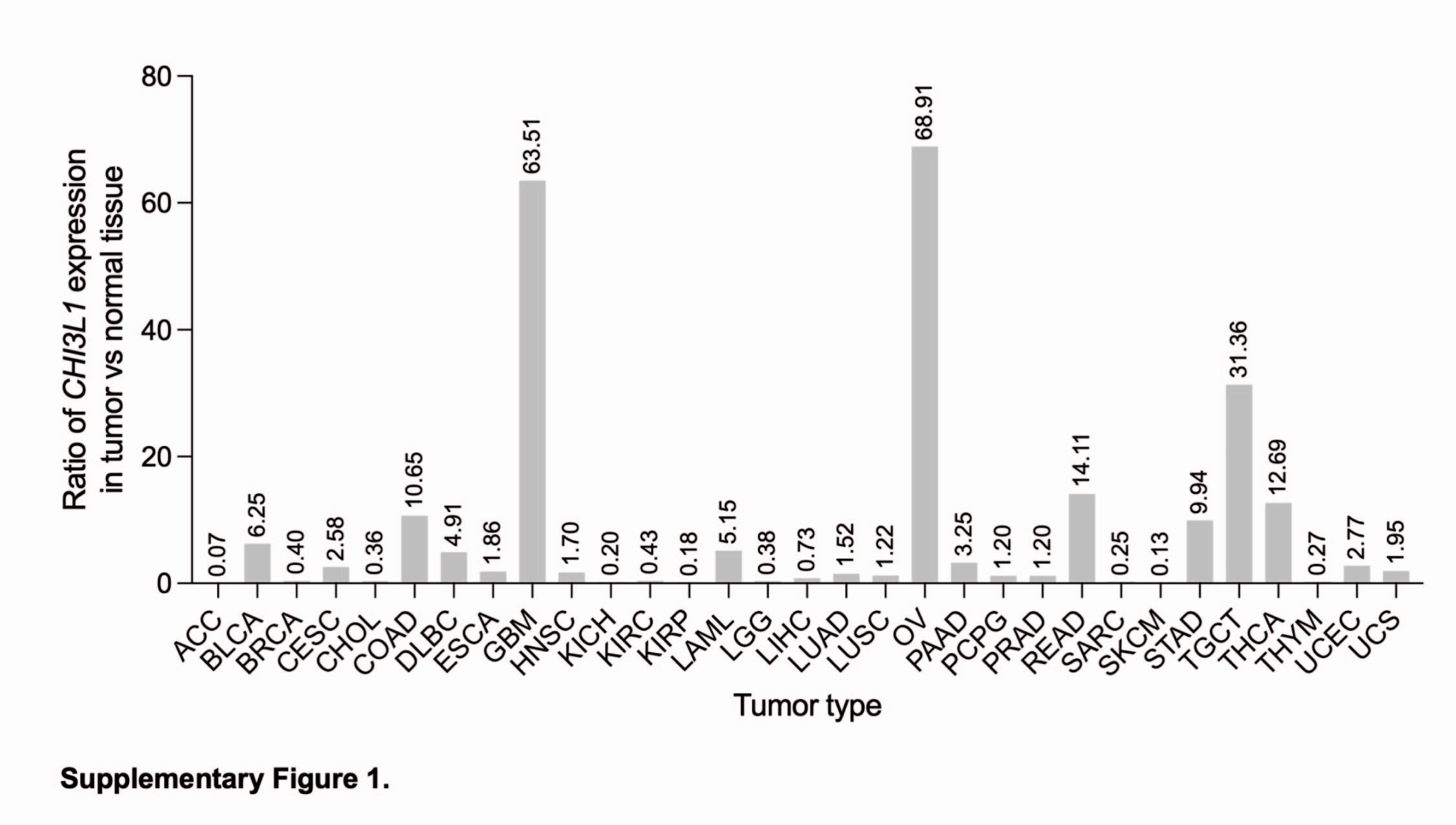
We determined CHI3L1 expression in glioma bulk and single-cell RNA sequencing (scRNAseq) datasets. Comparison of CHI3L1 expression in various tumors in the GEPIA database (17) shows that CHI3L1 expression is highest in GBM among 32 cancer types (Fig. 1A) with 63.5 times higher expression in GBM vs. normal brain (Supp. Figure 1). The quantitative PCR (RT-qPCR) analysis of CHI3L1 expression in the 76 gliomas dataset (18) shows upregulated expression of CHI3L1 in malignant (GBM) and benign gliomas (pilocytic astrocytoma) compared to normal brain tissue (Fig. 1B). Similarly, high CHI3L1 expression was detected in the bulk-RNAseq dataset of 31 gliomas dataset (19) (Fig. 1C). The analysis of the published scRNAseq dataset of 28 IDH1 wild-type GBMs shows that CHI3L1 was expressed predominantly by malignant cells, with some expression in glioma-associated macrophages (Fig. 1D-E). Interestingly, CHI3L1 expression positively correlated with the mesenchymal signature (MESlike1), the aggressive transcriptional subtype of GBM (Fig. 1E-F). We cross-checked the genes listed in the MESlike1 signature with the proteins correlated with CHI3L1 in the Brain Protein Atlas database (20,21). Out of 27 genes assigned to the MESlike1 signature, 25 correlated positively with CHI3L1 at a protein level in GBMs (Fig. 1G).
CHI3L1 depleted human glioma cells show altered expression of genes involved in extracellular matrix reorganization and cell adhesion
To evaluate the effect of CHI3L1 absence, CRISPR/Cas9 genome editing was applied and human glioma cells depleted of CHI3L1 (CHI3L1 KO) were generated (Fig. 2A). U87-MG cells exhibited high CHI3L1 protein level compared to normal astrocytes and were selected for further experiments (Fig. 2B). While WG9 cells showed highest CHI3L1 mRNA/protein levels, they failed to establish tumors in recipient mice (data not shown). We used the Cancer Dependency Map portal (22,23) and found U87-MG to have the highest gene effect (a negative parameter predicting the effect of gene knock-out on cell viability) for CHI3L1 depletion among the commonly used human glioma cell lines (Fig. 2C).
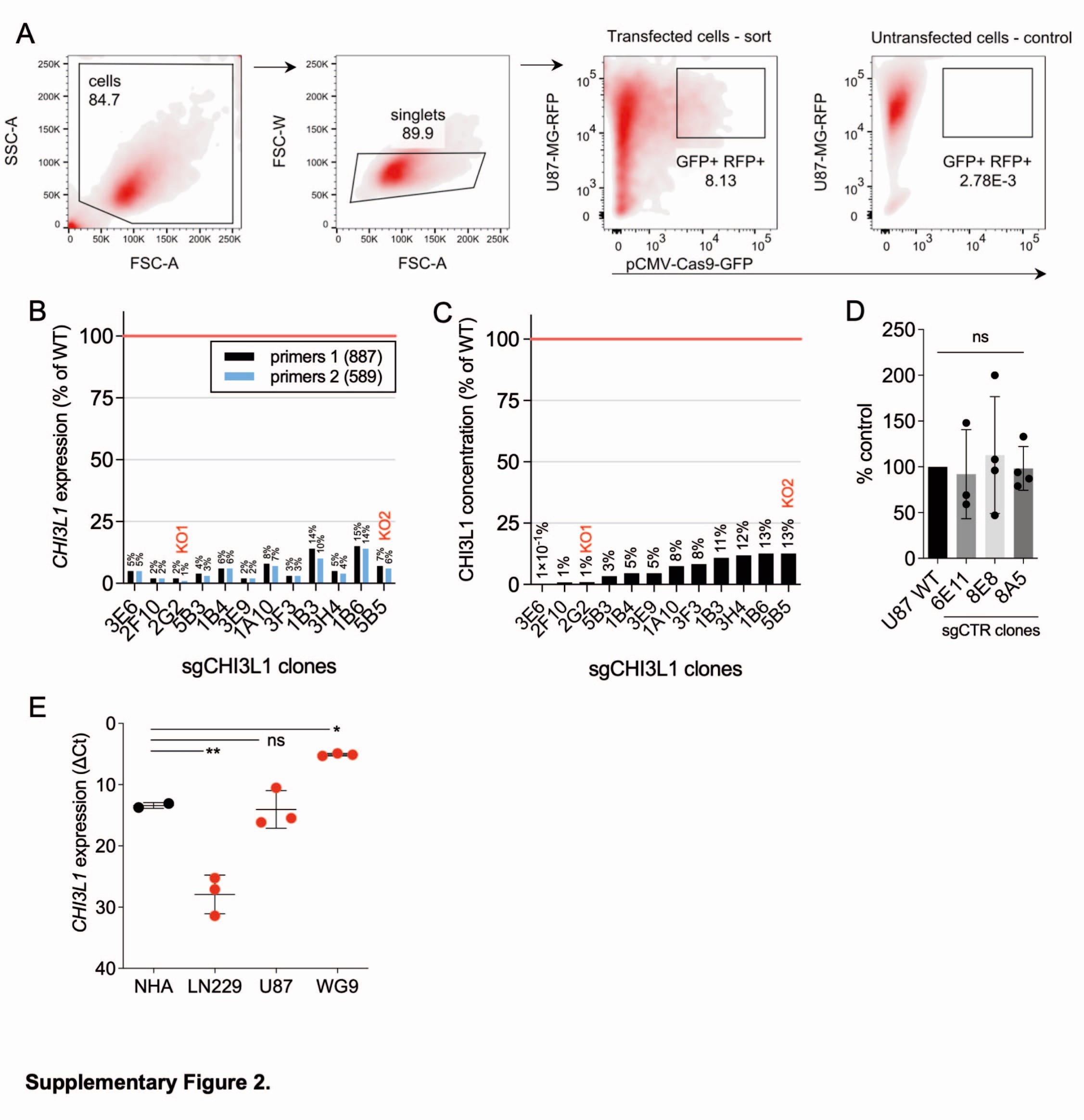
U87-MG cells were transfected with a commercial Cas9-GFP plasmid and validated guide RNAs specific for two sites in the CHI3L1 gene. Transfected cells (GFP+, Suppl. Figure 2A) were sorted individually and expanded clonally. Multiple independent clones were subjected to RT-qPCR and ELISA to evaluate efficacy of knockout (Suppl. Figure 2B-C). Two clones with the lowest CHI3L1 expression (KO1 and 2) were utilized further (Fig. 2D-E).
We performed RNAseq on wild-type (WT) and two CHI3L1 KO clones. A Venn diagram shows numbers of commonly up- or down-regulated genes among DEGs in KO1 and KO2 clones compared to WT cells (Fig. 2F). Around 50% of DEGs were affected in both KO cell lines. A Volcano plot shows DEGs in KO1 cells (Fig. 2G) and Reactome pathway enrichment analysis reveals that many genes down-regulated in CHI3L1 KO clones belong to ECM organization, migration, and invasion pathways (MMP2, MMP24, BMP1, PCOLCE, PCOLCE2, COL5A2, COL1A1, ADAMTS2, ADAMTS9, EFEMP1, TGFB3) (Fig. 2H).
Genes highly up-regulated in CHI3L1 KO cells included genes associated with formation and re-organization of ECM and cell-to-cell adhesion (FBLN1, PXDN, MMP1, PCDHB8) (Fig. 2G). Genes involved in PD-1 and IL-10 signaling were downregulated in CHI3L1 KO cells. CSF1 encoding macrophage colony stimulating factor or TGFB3 encoding transforming growth factor 3 were consistently downregulated in both clones (Fig. 2H). Some of the genes determined as mesenchymal-associated in single-cell database (24): ANXA1, EFEMP1, SPP1, NPC2, TNFRSF1A were most down-regulated in CHI3L1 KO compared to WT cells (Fig. 2G-H).
We cross-checked DEGs in CHI3L1 KO cells with the Brain Protein Atlas study (20,21) and found a positive correlation between CHI3L1 and proteins encoded by genes down-regulated in CHI3L1 KO cells (EFEMP1, ANXA1, NPC2, SERPINH1 and HLA-DRA) (Fig. 2I).
Altogether, CHI3L1 KO cells downregulated genes involved in ECM reorganization, invasion, and immunosuppression.
Glioblastoma cells depleted of CHI3L1 form smaller tumors in recipient mice
CHI3L1 KO glioma cells implanted intracranially into Foxn1nu mice developed significantly smaller tumors compared to WT controls as demonstrated by magnetic resonance imaging (MRI) 21 days post-implantation (Fig. 3A-B). Mice in CHI3L1 KO group gained weight during tumor growth, which is an indicator of a lesser tumor burden in this group (Fig. 3C). Tumor volume and body mass of tumor-bearing animals were negatively correlated (Fig. 3D).
Human CHI3L1 (hCHI3L1) was detected in blood serum of mice bearing WT tumors and observed to drop substantially in mice bearing CHI3L1 KO tumors (Fig. 3E, left). hCHI3L1 serum level positively correlated with tumor volume in the WT group (Fig. 3E, right). Mouse CHI3L1 was detectable in the serum of naive mice and increased in mice bearing WT tumors but not CHI3L1 KO tumors (Fig. 3F, left). mCHI3L1 levels did not correlate with tumor volume (Fig. 3F, right).
Importantly, diminished tumor growth in KO group was not due to reduced cell proliferation, as WT and CHI3L1 KO U87-MG cells had comparable proliferation rate in vitro (Fig. 3G-H). Adding the recombinant hCHI3L1 to culture media of CHI3L1 KO glioma cells did not increase CHI3L1 KO cell proliferation (Fig. 3I). Thus, extracellular CHI3L1 does not control glioblastoma proliferation in vitro. Notably, CRISPR/Cas9 manipulation did not affect proliferation of U87-MG cells (Suppl. Figure 2D).
Glioma-derived CHI3L1 enhances invasion and myeloid cell infiltration into the tumor
CHI3L1 KO cells showed lowered expression of MMP2, coding for metalloproteinase 2 (Fig. 4A). We performed gelatin zymography and found a significantly lower gelatinolytic activity in conditioned media from CHI3L1 KO1 glioma cells compared to WT cells (Fig. 4B). We and others had demonstrated that glioma invasion is strongly enhanced by microglial cells (25–27). Invasion of WT and CHI3L1 KO (KO1) glioma cells in the presence/absence of human HM-SV40 microglial cells was determined with a matrigel assay. We found reduced invasion of CHI3L1 KO cells in microglia presence (Fig. 4C).
Brain resident microglia and monocyte-derived macrophages accumulate and are reprogrammed by GBM to support tumor progression, immunosuppression, and resistance to therapy (28,29). One of the reprograming factors in GBM is SPP1 (osteopontin) (25). The expression of SPP1 was significantly down-regulated in CHI3L1 KO glioma cells (Fig. 4D), which was corroborated on protein level (Fig. 4E). We determined the infiltration of myeloid cells in intracranial gliomas by staining for IBA1, a pan-myeloid cell marker. We found the density of IBA1 + cells significantly decreased in CHI3L1 KO tumors compared to WT tumors (Fig. 4F-G).
Depletion of glioma-derived CHI3L1 results in normalization of tumor vasculature
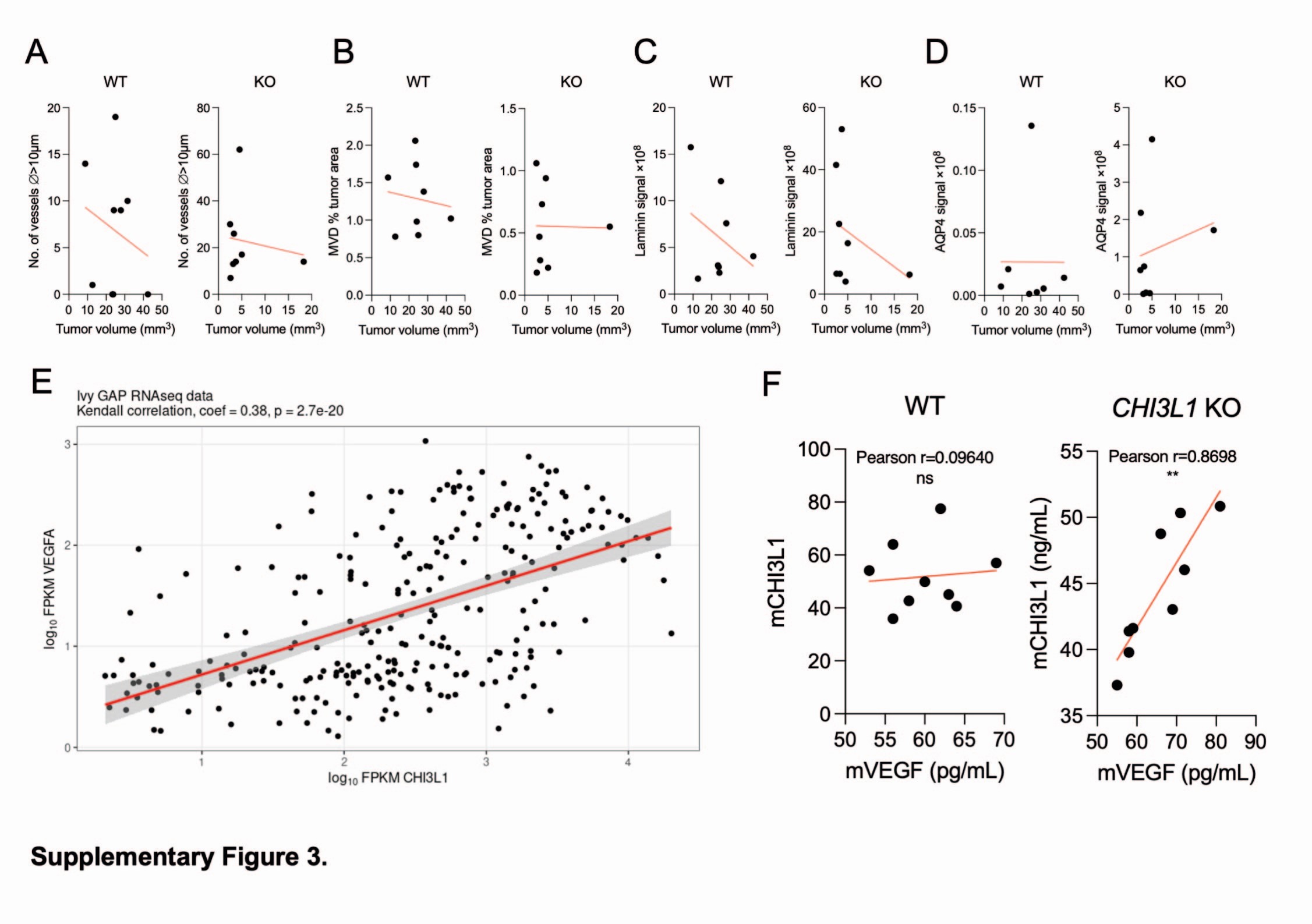
GBM is one of the most highly vascularized solid tumors and it is characterized by an extensive proliferation of microvessels (30,31), which leads to a high microvessel density (MVD). We compared the vasculature of WT and CHI3L1 KO U87-MG tumors by staining for von Willebrand Factor (vWF), an activated endothelium marker (Fig. 5A). The gross vasculature differed considerably in WT and CHI3L1 KO tumors; with more numerous non-capillary vessels (Fig. 5C left) and lesser MVD in the latter (Fig. 5C right). The number of vessels did not correlate with the tumor volume (Suppl. Figure 3A-D), suggesting that the increase in non-capillary vessels in CHI3L1 KO tumors was independent of tumor volume but resulted from CHI3L1 depletion instead.
We examined the structure of the blood vessel walls by staining tumor sections for laminin, an endothelium basement membrane marker (Fig. 5B). We observed that in CHI3L1 KO gliomas blood vessels were continuously lined with laminin in contrast to WT tumors. Laminin immunopositivity significantly increased in CHI3L1 KO gliomas vs. WT (Fig. 5D). The integrity of morphologically abnormal vessels was assessed using endothelium CD31 staining and 3D rendering. CD31 + vessels formed continuous tubes in CHI3L1 KO tumors in comparison to those in WT group (Fig. 5E). These findings show that in the absence of tumor-derived CHI3L1, blood vessels are better structured and likely more functional.
Coverage of blood vessels by astrocytic endfeet contributes to the blood-brain barrier by sealing endothelial cell junctions and controlling the transport of substances from blood (32). Astrocytes clear waste and excess of extracellular fluid from the perivascular space via a water channel aquaporin 4 (AQP4) (33), localized at the astrocytic endfeet. The expression of AQP4 was low in WT U87-MG gliomas but considerably increased in CHI3L1 KO gliomas (Fig. 5F and H). A linear regression analysis for CHI3L1 and AQP4 protein levels in the BPA database shows a negative correlation between the two proteins in GBM (Fig. 5I). Reduced staining for glial fibrillary acidic protein (GFAP) demonstrates that astrocytes were less activated in CHI3L1 KO tumors in comparison to WT controls (Fig. 5G and J). Aquaporin 4 was localized at the GFAP + cell protrusions covering blood vessels as demonstrated with double staining for AQP4 and GFAP (Fig. 5K). We propose that tumor-derived CHI3L1 promotes deregulation of the vascular network in gliomas by increasing the number of endothelial sprouts, altering the structure of vascular walls, and decreasing the coverage of blood vessels by astrocytes (Fig. 6).
{kind=link}
{kind=link}
{kind=link}